Курсовая на тему Общие свойства живых систем
Работа добавлена на сайт bukvasha.net: 2014-12-07Поможем написать учебную работу
Если у вас возникли сложности с курсовой, контрольной, дипломной, рефератом, отчетом по практике, научно-исследовательской и любой другой работой - мы готовы помочь.

Предоплата всего
от 25%

Подписываем
договор
Реферат
Общие свойства живых систем
1. Основные особенности и внутрисистемные связи живых систем
При всем своем многоплановом разнообразии живые системы имеют некоторые, хотя и немногочисленные, но неотъемлемые общие особенности, определяемые фундаментальными общими свойствами живого. Таких особенностей три: 1) наличие собственной программы развития системы, развертываемой на основе активно регулируемых информационных взаимосвязей ее с внешней средой, 2) иерархическая функционально-структурная организация и 3) высокая функционально-структурная сложность.
Наличие собственной программы развития — основополагающее свойство всех живых систем, поскольку оно определяет их главную общую черту — иерархическую функционально-структурную организацию, которая представляет собою высшую форму упорядоченности системы и служит источником ее антиэнтропичности. Именно иерархическая организация живых систем позволяет им достигать той характерной для них высокой функционально-структурной сложности, особенно на молекулярном уровне, которая и создает специфику живого на основе развития функционально необходимых внутренних информационных, материальных и энергетических связей живой системы и ее разносторонних контактов с внешней средой. Поэтому любое усложнение живой системы в итоге ведет к более разветвленной иерархии в ее целостной функционально-структурной организации. Оба эти процесса — общее усложнение системы и прогрессивное развитие иерархического характера ее организации — происходит на основе соответствующего усложнения собственной программы развития системы. В схематическом виде основные связи живой системы показаны на рис. 1.
Рис 1
2. Наличие собственной программы развития системы и способность к активному оперированию информацией
Всем живым системам любого уровня сложности, как организменным„ так и надорганизменным, совершенно независимо от их функционально-структурных особенностей, элементарной основы и всех других качеств, свойственно наличие определенной собственной программы развития, реализация которой обеспечивает сохранение системы в меняющихся условиях внешней среды.
Наличие собственной программы развития — важнейшая особенность живых систем, определяющая все их другие специфические свойства.
В зависимости от уровня функциональной организации живой системы собственная программа ее развития существенно изменяется по своей структуре, сложности и степени централизации. В связи с этим значительные различия существуют между собственными программами развития организменных и надорганизменных систем.
У организмов собственная программа развития имеет в своей основе генетическую программу, материализованную на молекулярном уровне в виде генома. Генетическая программа содержит конкретную генетическую информацию, необходимую для обеспечения онтогенетического развития организменной системы и ее самовоспроизведения в цикле развития вида. Эта программа в своей основе имеет: 1) закономерности кодирования, транскрипции и трансляции генетической информации; 2) закономерности репликации и сегрегации генетического материала; 3) закономерности самосборки первичных биологических структур, т. е. совокупности сложных биополимеров — нуклеиновых кислот и белков, взаимодействие между которыми обеспечивает возможность саморегуляции живых систем организменного уровня.
Собственная программа развития организма всегда является централизованной, однако формы и степень этой централизации различны в зависимости от уровня структурной агрегации организменной системы. Монобионтная программа развития всегда централизована на генетическом уровне, т.е. построена по принципу генетического моноцентризма; метабионтная программа лишена генетического моноцентризма и централизована уже на уровне межклеточных взаимодействий; а централизация ценометабионтной программы осуществляется на уровне взаимодействий многоклеточных суборганизменных структурных блоков, имеющих ранг метабионтов. Таким образом, с повышением уровня структурной агрегации организменной системы уровень централизации ее собственной программы развития закономерно удаляется от молекулярного уровня материализации генетической программы. Только у монобионтов эти уровни совпадают и генетическая программа представляет собою единственный элемент собственной программы развития организма, которая вследствие этого является генетически моноцентричной. В метабионтных и ценометабионтных организменных системах этот моноцентризм генетической программы сохраняется только на своем прежнем, монобионтном уровне, т.е. на уровне клетки.
Централизация собственной программы развития — важнейшая особенность информационной структуры организменных живых систем. Эта особенность, связанная с функционально-структурной целостностью и неделимостью организма, принципиально отличает любые организменные системы от надорганизменных.
В надорганизменных живых системах подобная централизованная программа развития отсутствует. Собственная программа развития надорганизменной системы формируется как интегральный результат взаимодействия некоторого множества ее централизованных подпрограмм, т.е. собственных программ развития тех организмов, которые образуют данную надорганизменную систему. Этот интегральный генофонд надорганиз-менной системы в конечном итоге образует столь же определенную программу ее развития на основе тех же принципов самовоспроизведения и саморегуляции, что и в случае отдельных организмов.
Способность к активному оперированию информацией столь же специфична для живых систем, как и само наличие собственной программы развития. Все биологические процессы связаны с той или иной формой активного оперирования с информацией и могут осуществляться лишь на основе постоянного поддержания в живой системе строго определенных параметров информационных потоков.
Говоря об активном оперировании информацией, мы хотели бы обратить внимание на тот факт, что неживые системы способны только к пассивному информационному обмену с окружающей их средой, который не включает в себя регулирование информационных потоков и специальную обработку поступающей в систему информации. Для живых систем характерно, напротив, 1) постоянное активное регулирование параметров двусторонних информационных потоков, связывающих систему с внешней средой, и 2) постоянная обработка, накопление и хранение поступающей в систему информации, что составляет необходимое условие реализации собственной программы развития живой системы.
Таким образом, развертывание собственной программы развития живой системы состоит в постоянном оперировании информацией, без чего эта программа не может быть реализована. Как некоторый конечный по объему комплекс информации собственная программа развития сама по себе еще не является достаточной для управления всей совокупностью жизненных процессов, осуществляемых живой системой. Информационный комплекс программы может быть приведен в действие лишь на основе постоянного притока информации извне, т. е. во взаимодействии с информационным полем окружающей среды, что представляет собою один из аспектов взаимосвязи живой системы с окружающей средой.
Информационный баланс живой системы всегда является положительным. Это правило не имеет исключений и составляет основу понимания жизни как процесса, связанного с уменьшением энтропии в живых системах при одновременном возрастании ее в окружающей их среде. Антиэнтропичность живых систем выражается, в частности, в их роли как концентраторов информации, своеобразных «информационных сгустков».
Способность живых систем к концентрированию информации обусловлена их иерархической структурой и их высокой функционально-структурной сложностью.
Наличие собственной программы развития — изначальное свойство живых систем. Именно возникновение программы развития, представляющей собою необходимое условие самоповторения системы, привело к появлению организмов как относительно стабильных в качественном отношении дискретных единиц живого и явилось исходным пунктом собственно биологической эволюции. В этом смысле эволюция организмов есть эволюция их собственных программ развития.
Пространственно-временная дискретность живого, находящая свое выражение в существовании организмов, предъявляет весьма жесткие требования к структуре и качеству информационного потока как в рамках онтогенеза, где этот поток определяет все элементы функционально-структурной специфики развивающейся организменной системы и порядок выражения отдельных морфопоэтических факторов, так и в филогенетическом, эволюционном плане, где от упорядоченности информационного потока зависит точность передачи всех свойств живой системы в чреде поколений и, соответственно, мера преемственности результатов эволюционного процесса. Эти строго определенные параметры информационного потока, создающего достаточную основу для управления процессами онтогенеза и филогенеза, и обеспечиваются конкретной пограммой развития, содержащейся в материализованной форме в живых системах организмен-ного уровня. Естественно поэтому, что без собственной программы развития были бы невозможны ни самовоспроизводство живых систем, ни их эволюция.
В необозримом многообразии живых систем можно различать разные уровни сложности собственной программы развития системы и различные формы записи ее элементов, но это — уже второстепенные детали. Главное и общее состоит в том, что собственная программа развития свойственна всем живым системам и всегда функционирует во взаимосвязи с информационным полем окружающей среды, причем живая система непрерывно потребляет информацию извне и активно оперирует ею. Заметим, что под оперированием информацией следует понимать не только традиционно включаемые в это понятие действия органов чувств или обмен химическими сигналами внутри организма, но также и усвоение различных веществ в процессе питания или акцептора электронов в процессе дыхания. Действительно, в этом смысле вряд ли можно видеть принципиальную разницу между усвоением молекул любого вещества в кишечнике и восприятием мышечной клеткой медиатора, выделяемого аксоном в синаптическую щель. В обоих случаях происходит усвоение информации, содержащейся в структуре конкретной молекулы, только в первом случае эта информация подвергается организмом длительной переработке, тогда как во втором — анализ ее совершается в доли секунды.
3. Иерархическая функционально-структурная организация
Все живые системы отличаются высокой упорядоченностью, которая проявляется в их целостной, иерархической функционально-структурной организации и находит свое выражение во взаимообусловленности всех отдельных элементов системы и их подчиненности ее общему функционально-структурному плану. И хотя эта иерархичность организации составляет общее свойство любых систем, в случае живых систем она приобретает совершенно особое значение, поскольку специфика этих систем создается благодаря их чрезвычайно высокой функционально-структурной сложности, которая требует исключительно тонкого согласования всех подсистем в пределах системы при весьма динамичном управлении всей многогранной совокупностью одновременно происходящих процессов, связанных с развитием этих подсистем.
Поэтому обеспечение экологически целесообразного интегрального результата деятельности живой системы, необходимого для ее сохранения и адаптивного развития, в любом случае возможно только на основе ее строго иерархической общей организации, при которой задача управления системой принципиально упрощается за счет того, что отдельные элементы этой сложной задачи могут решаться автономно и последовательно на отдельных иерархически соподчиненных уровнях системы. Только на основе этой иерархии процессов и структур в живой системе и становится возможным как фактически существующее в ней полное взаимосоответствие внутренних потоков информации, энергии и вещества, так и экологически целесообразное развитие ее информационных, энергетических и материальных связей с внешней средой. Иначе говоря, саморегуляция системы подобного уровня сложности может осуществляться только на основе ее строго иерархической, целостной организации.
Иерархическая структура как высшая форма упорядоченности свойственна живым системам на всех уровнях организации, начиная с субмолекулярного уровня.
На субмолекулярном и молекулярном уровнях примерами иерархической организации живых систем могут служить любые биологические макромолекулы, и прежде всего гигантские молекулы нуклеиновых кислот и белков с их сложными, разноуровневыми эволюционно обусловленными субъединицами, архитектура которых непосредственно определена многогранными совокупностями внутримолекулярных связей различной природы. Не повторяя общеизвестных истин химии нуклеиновых кислот и белков, напомним лишь, что они организованы именно на основе жесткой многоуровневой функционально-структурной иерархии определенных элементарных блоков разной сложности. В этой связи рассмотрим несколько подробнее только один пример — структурную организацию глобулярных белков.
Пространственная структура глобулярного белка представляет собою результат сложной многоуровневой иерархической организации первичных компонентов — аминокислот, причем в первичной структуре, т. е. «в аминокислотной последовательности заключена полная информация... Поэтому между всеми уровнями имеется взаимосвязь, так что элементы более низких уровней определяют элементы более высоких уровней».
Первый структурный уровень, или 'первичная структура белка, представляет собою последовательность аминокислотных остатков в полипептидной цепи, связанных между собою ковалентными связями. Второй структурный уровень, или вторичная структура белка,— это упорядоченное, энергетически предпочтительное спиральное или зигзагообразное расположение полипептидных цепей, стабилизированное за счет водородных связей. Третий структурный уровень, или третичная структура белка, определяет способ укладки полипептидной цепи с образованием очень компактной, плотно упакованной структуры — глобулы. Третичная, структура стабилизирована связями разнообразной природы. Четвертый структурный уровень, или четвертичная структура белка, представляет собою функционально целостные макроагрегаты из двух или большего числа полипептидных цепей — протомеров, т. е. так называемый олигомерный белок.
Важно подчеркнуть, что «в аминокислотной последовательности полипептидной цепи олигомерного белка закодирована информация не только о его третичной структуре, но также и о геометрической форме контактных участков, с которыми соединяются контактные участки других специфических протомеров. Таким образом, информация, содержащаяся в аминокислотной последовательности, может реализоваться на нескольких уровнях структурной организации — вторичном, третичном и четвертичном». Это ясно указывает на иерархический характер организации белка.
Общим для белка «свойством, проявляющимся на всех уровнях организации выше аминокислотного, является симметрия... Наличие сим^ метрии и иерархии уровней белковых структур позволяет более ясно представить некоторые аспекты сложнейшей проблемы пространственной организации белковых молекул».
На надмолекулярном суборганизменном уровне иерархичность организации живых систем общеизвестна и достаточно демонстративна: все названные структуры представляют собою конкретные подсистемы организма и как таковые принципиально возможны только в рамках целостной функционально-структурной системы организменного уровня.
На организменном уровне иерархичность организации столь же очевидна, как и на суборганизменных уровнях: иерархическая соподчиненность всех функционально-структурных элементов организма составляет одну из его главных особенностей и характеризует его как неделимую единицу жизни.
На надорганизменном уровне иерархическая организация живых систем имеет место всегда, хотя в разных случаях может быть выражена в различной степени.
В структурно неупорядоченных временных надорганизменных системах эта иерархия может проявляться относительно слабо, примером чего могут служить хотя бы миграционные стаи мелких птиц, в частности многих воробьинообразных. Стая скворцов не является структурно упорядоченной в том смысле, что расположение птиц относительно друг друга в ней не подчиняется какому-либо твердому закону. В то же время поведение каждой птицы в летящей стае подчинено поведению стаи, в чем проявляется иерархическая, системная структура стаи.
Общие свойства живых систем
1. Основные особенности и внутрисистемные связи живых систем
При всем своем многоплановом разнообразии живые системы имеют некоторые, хотя и немногочисленные, но неотъемлемые общие особенности, определяемые фундаментальными общими свойствами живого. Таких особенностей три: 1) наличие собственной программы развития системы, развертываемой на основе активно регулируемых информационных взаимосвязей ее с внешней средой, 2) иерархическая функционально-структурная организация и 3) высокая функционально-структурная сложность.
Наличие собственной программы развития — основополагающее свойство всех живых систем, поскольку оно определяет их главную общую черту — иерархическую функционально-структурную организацию, которая представляет собою высшую форму упорядоченности системы и служит источником ее антиэнтропичности. Именно иерархическая организация живых систем позволяет им достигать той характерной для них высокой функционально-структурной сложности, особенно на молекулярном уровне, которая и создает специфику живого на основе развития функционально необходимых внутренних информационных, материальных и энергетических связей живой системы и ее разносторонних контактов с внешней средой. Поэтому любое усложнение живой системы в итоге ведет к более разветвленной иерархии в ее целостной функционально-структурной организации. Оба эти процесса — общее усложнение системы и прогрессивное развитие иерархического характера ее организации — происходит на основе соответствующего усложнения собственной программы развития системы. В схематическом виде основные связи живой системы показаны на рис. 1.
| |
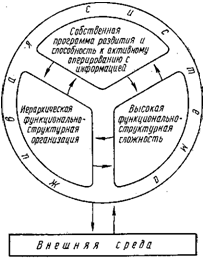 |
Рис 1
2. Наличие собственной программы развития системы и способность к активному оперированию информацией
Всем живым системам любого уровня сложности, как организменным„ так и надорганизменным, совершенно независимо от их функционально-структурных особенностей, элементарной основы и всех других качеств, свойственно наличие определенной собственной программы развития, реализация которой обеспечивает сохранение системы в меняющихся условиях внешней среды.
Наличие собственной программы развития — важнейшая особенность живых систем, определяющая все их другие специфические свойства.
В зависимости от уровня функциональной организации живой системы собственная программа ее развития существенно изменяется по своей структуре, сложности и степени централизации. В связи с этим значительные различия существуют между собственными программами развития организменных и надорганизменных систем.
У организмов собственная программа развития имеет в своей основе генетическую программу, материализованную на молекулярном уровне в виде генома. Генетическая программа содержит конкретную генетическую информацию, необходимую для обеспечения онтогенетического развития организменной системы и ее самовоспроизведения в цикле развития вида. Эта программа в своей основе имеет: 1) закономерности кодирования, транскрипции и трансляции генетической информации; 2) закономерности репликации и сегрегации генетического материала; 3) закономерности самосборки первичных биологических структур, т. е. совокупности сложных биополимеров — нуклеиновых кислот и белков, взаимодействие между которыми обеспечивает возможность саморегуляции живых систем организменного уровня.
Собственная программа развития организма всегда является централизованной, однако формы и степень этой централизации различны в зависимости от уровня структурной агрегации организменной системы. Монобионтная программа развития всегда централизована на генетическом уровне, т.е. построена по принципу генетического моноцентризма; метабионтная программа лишена генетического моноцентризма и централизована уже на уровне межклеточных взаимодействий; а централизация ценометабионтной программы осуществляется на уровне взаимодействий многоклеточных суборганизменных структурных блоков, имеющих ранг метабионтов. Таким образом, с повышением уровня структурной агрегации организменной системы уровень централизации ее собственной программы развития закономерно удаляется от молекулярного уровня материализации генетической программы. Только у монобионтов эти уровни совпадают и генетическая программа представляет собою единственный элемент собственной программы развития организма, которая вследствие этого является генетически моноцентричной. В метабионтных и ценометабионтных организменных системах этот моноцентризм генетической программы сохраняется только на своем прежнем, монобионтном уровне, т.е. на уровне клетки.
Централизация собственной программы развития — важнейшая особенность информационной структуры организменных живых систем. Эта особенность, связанная с функционально-структурной целостностью и неделимостью организма, принципиально отличает любые организменные системы от надорганизменных.
В надорганизменных живых системах подобная централизованная программа развития отсутствует. Собственная программа развития надорганизменной системы формируется как интегральный результат взаимодействия некоторого множества ее централизованных подпрограмм, т.е. собственных программ развития тех организмов, которые образуют данную надорганизменную систему. Этот интегральный генофонд надорганиз-менной системы в конечном итоге образует столь же определенную программу ее развития на основе тех же принципов самовоспроизведения и саморегуляции, что и в случае отдельных организмов.
Способность к активному оперированию информацией столь же специфична для живых систем, как и само наличие собственной программы развития. Все биологические процессы связаны с той или иной формой активного оперирования с информацией и могут осуществляться лишь на основе постоянного поддержания в живой системе строго определенных параметров информационных потоков.
Говоря об активном оперировании информацией, мы хотели бы обратить внимание на тот факт, что неживые системы способны только к пассивному информационному обмену с окружающей их средой, который не включает в себя регулирование информационных потоков и специальную обработку поступающей в систему информации. Для живых систем характерно, напротив, 1) постоянное активное регулирование параметров двусторонних информационных потоков, связывающих систему с внешней средой, и 2) постоянная обработка, накопление и хранение поступающей в систему информации, что составляет необходимое условие реализации собственной программы развития живой системы.
Таким образом, развертывание собственной программы развития живой системы состоит в постоянном оперировании информацией, без чего эта программа не может быть реализована. Как некоторый конечный по объему комплекс информации собственная программа развития сама по себе еще не является достаточной для управления всей совокупностью жизненных процессов, осуществляемых живой системой. Информационный комплекс программы может быть приведен в действие лишь на основе постоянного притока информации извне, т. е. во взаимодействии с информационным полем окружающей среды, что представляет собою один из аспектов взаимосвязи живой системы с окружающей средой.
Информационный баланс живой системы всегда является положительным. Это правило не имеет исключений и составляет основу понимания жизни как процесса, связанного с уменьшением энтропии в живых системах при одновременном возрастании ее в окружающей их среде. Антиэнтропичность живых систем выражается, в частности, в их роли как концентраторов информации, своеобразных «информационных сгустков».
Способность живых систем к концентрированию информации обусловлена их иерархической структурой и их высокой функционально-структурной сложностью.
Наличие собственной программы развития — изначальное свойство живых систем. Именно возникновение программы развития, представляющей собою необходимое условие самоповторения системы, привело к появлению организмов как относительно стабильных в качественном отношении дискретных единиц живого и явилось исходным пунктом собственно биологической эволюции. В этом смысле эволюция организмов есть эволюция их собственных программ развития.
Пространственно-временная дискретность живого, находящая свое выражение в существовании организмов, предъявляет весьма жесткие требования к структуре и качеству информационного потока как в рамках онтогенеза, где этот поток определяет все элементы функционально-структурной специфики развивающейся организменной системы и порядок выражения отдельных морфопоэтических факторов, так и в филогенетическом, эволюционном плане, где от упорядоченности информационного потока зависит точность передачи всех свойств живой системы в чреде поколений и, соответственно, мера преемственности результатов эволюционного процесса. Эти строго определенные параметры информационного потока, создающего достаточную основу для управления процессами онтогенеза и филогенеза, и обеспечиваются конкретной пограммой развития, содержащейся в материализованной форме в живых системах организмен-ного уровня. Естественно поэтому, что без собственной программы развития были бы невозможны ни самовоспроизводство живых систем, ни их эволюция.
В необозримом многообразии живых систем можно различать разные уровни сложности собственной программы развития системы и различные формы записи ее элементов, но это — уже второстепенные детали. Главное и общее состоит в том, что собственная программа развития свойственна всем живым системам и всегда функционирует во взаимосвязи с информационным полем окружающей среды, причем живая система непрерывно потребляет информацию извне и активно оперирует ею. Заметим, что под оперированием информацией следует понимать не только традиционно включаемые в это понятие действия органов чувств или обмен химическими сигналами внутри организма, но также и усвоение различных веществ в процессе питания или акцептора электронов в процессе дыхания. Действительно, в этом смысле вряд ли можно видеть принципиальную разницу между усвоением молекул любого вещества в кишечнике и восприятием мышечной клеткой медиатора, выделяемого аксоном в синаптическую щель. В обоих случаях происходит усвоение информации, содержащейся в структуре конкретной молекулы, только в первом случае эта информация подвергается организмом длительной переработке, тогда как во втором — анализ ее совершается в доли секунды.
3. Иерархическая функционально-структурная организация
Все живые системы отличаются высокой упорядоченностью, которая проявляется в их целостной, иерархической функционально-структурной организации и находит свое выражение во взаимообусловленности всех отдельных элементов системы и их подчиненности ее общему функционально-структурному плану. И хотя эта иерархичность организации составляет общее свойство любых систем, в случае живых систем она приобретает совершенно особое значение, поскольку специфика этих систем создается благодаря их чрезвычайно высокой функционально-структурной сложности, которая требует исключительно тонкого согласования всех подсистем в пределах системы при весьма динамичном управлении всей многогранной совокупностью одновременно происходящих процессов, связанных с развитием этих подсистем.
Поэтому обеспечение экологически целесообразного интегрального результата деятельности живой системы, необходимого для ее сохранения и адаптивного развития, в любом случае возможно только на основе ее строго иерархической общей организации, при которой задача управления системой принципиально упрощается за счет того, что отдельные элементы этой сложной задачи могут решаться автономно и последовательно на отдельных иерархически соподчиненных уровнях системы. Только на основе этой иерархии процессов и структур в живой системе и становится возможным как фактически существующее в ней полное взаимосоответствие внутренних потоков информации, энергии и вещества, так и экологически целесообразное развитие ее информационных, энергетических и материальных связей с внешней средой. Иначе говоря, саморегуляция системы подобного уровня сложности может осуществляться только на основе ее строго иерархической, целостной организации.
Иерархическая структура как высшая форма упорядоченности свойственна живым системам на всех уровнях организации, начиная с субмолекулярного уровня.
На субмолекулярном и молекулярном уровнях примерами иерархической организации живых систем могут служить любые биологические макромолекулы, и прежде всего гигантские молекулы нуклеиновых кислот и белков с их сложными, разноуровневыми эволюционно обусловленными субъединицами, архитектура которых непосредственно определена многогранными совокупностями внутримолекулярных связей различной природы. Не повторяя общеизвестных истин химии нуклеиновых кислот и белков, напомним лишь, что они организованы именно на основе жесткой многоуровневой функционально-структурной иерархии определенных элементарных блоков разной сложности. В этой связи рассмотрим несколько подробнее только один пример — структурную организацию глобулярных белков.
Пространственная структура глобулярного белка представляет собою результат сложной многоуровневой иерархической организации первичных компонентов — аминокислот, причем в первичной структуре, т. е. «в аминокислотной последовательности заключена полная информация... Поэтому между всеми уровнями имеется взаимосвязь, так что элементы более низких уровней определяют элементы более высоких уровней».
Первый структурный уровень, или 'первичная структура белка, представляет собою последовательность аминокислотных остатков в полипептидной цепи, связанных между собою ковалентными связями. Второй структурный уровень, или вторичная структура белка,— это упорядоченное, энергетически предпочтительное спиральное или зигзагообразное расположение полипептидных цепей, стабилизированное за счет водородных связей. Третий структурный уровень, или третичная структура белка, определяет способ укладки полипептидной цепи с образованием очень компактной, плотно упакованной структуры — глобулы. Третичная, структура стабилизирована связями разнообразной природы. Четвертый структурный уровень, или четвертичная структура белка, представляет собою функционально целостные макроагрегаты из двух или большего числа полипептидных цепей — протомеров, т. е. так называемый олигомерный белок.
Важно подчеркнуть, что «в аминокислотной последовательности полипептидной цепи олигомерного белка закодирована информация не только о его третичной структуре, но также и о геометрической форме контактных участков, с которыми соединяются контактные участки других специфических протомеров. Таким образом, информация, содержащаяся в аминокислотной последовательности, может реализоваться на нескольких уровнях структурной организации — вторичном, третичном и четвертичном». Это ясно указывает на иерархический характер организации белка.
Общим для белка «свойством, проявляющимся на всех уровнях организации выше аминокислотного, является симметрия... Наличие сим^ метрии и иерархии уровней белковых структур позволяет более ясно представить некоторые аспекты сложнейшей проблемы пространственной организации белковых молекул».
На надмолекулярном суборганизменном уровне иерархичность организации живых систем общеизвестна и достаточно демонстративна: все названные структуры представляют собою конкретные подсистемы организма и как таковые принципиально возможны только в рамках целостной функционально-структурной системы организменного уровня.
На организменном уровне иерархичность организации столь же очевидна, как и на суборганизменных уровнях: иерархическая соподчиненность всех функционально-структурных элементов организма составляет одну из его главных особенностей и характеризует его как неделимую единицу жизни.
На надорганизменном уровне иерархическая организация живых систем имеет место всегда, хотя в разных случаях может быть выражена в различной степени.
В структурно неупорядоченных временных надорганизменных системах эта иерархия может проявляться относительно слабо, примером чего могут служить хотя бы миграционные стаи мелких птиц, в частности многих воробьинообразных. Стая скворцов не является структурно упорядоченной в том смысле, что расположение птиц относительно друг друга в ней не подчиняется какому-либо твердому закону. В то же время поведение каждой птицы в летящей стае подчинено поведению стаи, в чем проявляется иерархическая, системная структура стаи.
Напротив, в структурно упорядоченных постоянных надорганизменных системах иерархическая структура выражена наиболее четко. В качестве примера можно указать ярусную структуру лиственного леса лесостепной зоны СССР, состоящего из основного — древесного яруса, ниже которого закономерно располагаются другие ярусы — кустарниковый, травяно-кустарничковый, моховой и т. д.; столь же выражена эта ярусность и по глубине укоренения растений. Такая ярусная иерархия над организменной системы леса оптимизирует распределение «сфер влияния» между представителями отдельных ярусов и тем самым ведет к наиболее полному освоению системой жизненных ресурсов.
Антиэнтропичность живых систем непосредственно связана именно с их упорядоченностью и проявляется в том, что свойственный им уровень энтропии ниже возможного максимума, соответствующего термодинамически равновесному состоянию системы. Поддержание такого состояния, когда уровень энтропии системы отличается от максимума, возможно в двух случаях:
при положительном энергобалансе системы, когда потребляемая ею из окружающей среды энергия больше выделяемой, что, в частности, соответствует метаболически активному состоянию живой системы; в плане общей теории систем это состояние должно быть определено как состояние открытой системы;
при отсутствии такого положительного энергобаланса, если система способна сохранять определенный, отличный от максимума, уровень энтропии только за счет своих структурных антиэнтропических свойств, препятствующих увеличению энтропии при отсутствии активно осуществляемого организмом материального обмена с окружающей средой, что, в частности, соответствует метаболически неактивному состоянию живой системы, т. е. состоянию полного анабиоза; с позиций общей теории систем такое состояние отвечает состоянию закрытой системы.
Оба эти состояния, в частности структурные антиэнтропические свойства закрытых живых систем. Важно подчеркнуть, что антиэнтропичность не есть какое-то самостоятельное свойство живых систем; она представляет собою лишь следствие реально существующей высокой упорядоченности этих систем.
4. Высокая функционально-структурная сложность
Общим свойством всех живых систем, независимо от общего уровня их организации, является их высокая функционально-структурная сложность.
При сколь угодно простой макроорганизации живые системы неизменно остаются высокосложными за счет своей специфической микроорганизации: при любой сложности макроорганизации живой системы ее интегральная сложность всегда определяется ее микроорганизацией на молекулярном и субмолекулярном уровнях. Принимая во внимание все информационные, энергетические и материальные аспекты молекулярной и субмолекулярной организации живых систем, следует признать, что преобладание этой молекулярной составляющей в интегральной сложности живой системы в любом случае столь значительно, что совокупность всех ее остальных, т.е. надмолекулярных, компонентов, бесспорно, имеет лишь подчиненное значение. В этом смысле можно сказать, что по степени суммарной сложности шимпанзе меньше отличается от вируса табачной мозаики, чем молекулярная организация шимпанзе от ее надмолекулярной организации.
Основанием для такого заключения служат общеизвестные истины молекулярной биологии. Представление о степени информационной и функционально-структурной сложности живых систем на молекулярном уровне дают хотя бы общеизвестные особенности генетического кода организмов. Достаточно напомнить, например, что один ген, обеспечивающий кодирование только одного полипептида, содержит примерно 1,5 X. X 103 нуклеотидных пар в ДНК, а палиндромные последовательности в ДНК эукариот, представляющие собою области узнавания для рестриктаз, содержат до 1,2 • 103 нуклеотидных пар. В свете подобных цифр, число которых можно было бы увеличить, высокая информационная и функционально-структурная сложность молекулярной организации живых систем представляется достаточно очевидной.
Естественно, что пример с шимпанзе не претендует на абсолютную точность; однако он, несомненно, верно отражает нашу общую мысль о решающей роли молекулярных особенностей в определении общего интегрального уровня сложности живой системы. Это, разумеется не дает нам права сбрасывать со счетов элементы сложности надмолекулярных уровней; суть дела состоит лишь в том, что совокупность этих надмолекулярных элементов по своему относительному значению в суммарном выражении сложности живой системы любого ранга явно уступает молекулярной составляющей.
Такое положение вполне соответствует тому факту, что все происходящие в организмах важнейшие жизненные процессы, связанные с изменениями информационных, энергетических и материальных состояний живой системы, структурно обеспечены на молекулярном и субмолекулярном уровнях. Именно это обстоятельство делает возможным, в частности, существование неклеточных живых систем, т. е. вирусов, у которых вся функционально-структурная организация ограничена уровнем макромолекул и сравнительно несложных надмолекулярных агрегатов. В этом одно из важнейших свидетельств особой весомости молекулярного уровня в многоуровневой целостной конструкции живых систем любой сложности.
Высокая сложность живых систем, основанная на их упорядоченной, строго иерархической функционально-структурной организации, в сочетании с программным управлением всеми протекающими в рамках системы процессами информационного, энергетического и материального плана, и создает ту глубокую специфику живых систем, которая поднимает их на более высокий, качественно новый уровень организации, принципиально отличающий их от объектов неживой природы.
Принимая во внимание все сказанное, мы можем заметить, что с принципиальной стороны жизнь как общее явление во Вселенной представляет собою феномен скорее кибернетического, чем субстратного плана. В конечном итоге не белки и не нуклеиновые кислоты как конкретные полимерные субстраты, а прежде всего их свойства как накопителей и преобразователей информации явились необходимой основой формирования антиэнтропических самоповторяющихся и эволюционирующих, т. е. живых, систем: при наличии этих информационных свойств на какой-либо иной субстратной основе явление жизни в принципе могло бы развиться с таким же успехом.
Понятие живой системы относится к объектам весьма различного уровня сложности. Содержание этого понятия может быть определено лишь в терминах биокибернетики и в соответствии с идеей системной организации, на основе учета некоторых общих принципов функционирования живых систем, в частности в аспектах связей, контроля и управления. В соответствии с этим в общем определении живой системы не могут найти отражения конкретно-биологические особенности, характеризующие специфику тех или иных частных систем любого уровня: такие особенности могут быть учтены лишь в определениях живых систем конкретного уровня сложности, например в определении открытой и закрытой живых систем.
Принимая во внимание большое разнообразие уровней организации известных в настоящее время живых систем и существующие между ними различия в формах их связей с окружающей их средой, общее определение живой системы можно сформулировать в следующем виде.
Живая система — иерархически организованная сложная система, имеющая собственную программу развития, реализация которой обеспечивает сохранение системы на основе поддержания в ней определенного, отличного, от максимума уровня энтропии. Это определение подчеркивает, таким образом, все три основные особенности живой системы.
Биологи, несомненно, обратили внимание на то, что в приведенном выше определении живой системы отсутствуют указания на традиционно выделяемые свойства живого. Это объясняется тем, что эти «традиционные» свойства в большинстве своем могут относиться не ко всем, а только к открытым живым системам, с которыми их и было принято связывать. Иначе говоря, прежние определения живого, содержащие эти «традиционные» элементы, оставляли за пределами живого закрытые живые системы, соответствующие состоянию полного анабиоза. Естественно, что наше общее определение, относящееся ко всем без исключения живым системам, не может содержать в себе элементы, приемлемые только для открытых систем.
Как видно из приведенного определения, оно не содержит указания на нуклеиново-белковый характер субстратов живых систем. Это обусловлено тем фактом, что все три свойственные живым системам особенности принципиально могут быть реализованы не только на основе нуклеиново-белковых субстратов, но и на иной химической основе, в связи с чем этот нуклеиново-белковый вариант организации живой системы не может найти отражения в ее общем определении.
Попытки обосновать общее определение живой системы исходя из нуклеиново-белковой природы земной жизни закономерно приводят к ограничительному толкованию этого понятия. Одним из примеров такого подхода может служить хотя бы следующее определение: «Живыми называются такие системы, которые обладают нуклеиновыми кислотами и белками и способны сами синтезировать эти вещества. Это определение неприложимо к древнейшим ступеням возникновения жизни, а также к существующим, возможно, внеземным живым системам, которые могут быть устроены иначе». Как видно из этого текста, подобное определение требует немедленных оговорок по поводу ограниченности сферы его применения. Кроме уже содержащихся в этом тексте ограничений, авторам вышеприведенного определения приходится считать, что вирусы — это объекты, «состоящие из нуклеиновых кислот и белков, но не являющиеся живыми», что противоречит их же собственному определению живой системы. Таким образом, попытки определения живой системы на основе констатации нуклеиново-белковой природы земных организмов исходят из предпосылок частного характера, т.е. страдают геоцентризмом и в этом смысле отражают пройденный этап развития биологии.
Наше определение не содержит также и упоминания р метаболической деятельности живых систем, поскольку она не свойственна закрытым живым системам.
Далее, в нашем определении ничего не сказано о способности живых систем к самовоспроизведению. Это связано с тем, что не все живые системы являются самовоспроизводящимися. Достаточно вспомнить многие бесплодные касты насекомых, например рабочих пчел и термитов и т.п.: все они представляют собою примеры несамовоспроизводящихся живых систем. То же можно сказать и о многих живых системах надорганизмеииого уровня, к которым понятие «самовоспроизведения» неприменимо, каковы, в частности, временные, миграционные стаи многих животных — различных рыб, птиц и др.
Наше определение не содержит и указаний на способность живых систем к эволюции, поскольку эволюционировать способны не все, а только метаболически активные самовоспроизводящиеся живые системы. Что же касается закрытых живых систем, представленных организмами, находящимися в состоянии полного анабиоза, то они не способны ни к самовоспроизводству, ни к эволюции, поскольку существуют в форме нефункционирующей структурной системы организма. Эволюционное развитие адаптации, обеспечивающих биологическую допустимость анабиотических состояний, происходит лишь за счет тех или иных генетических изменений, возникающих в метаболически активных фазах онтогенеза, тогда как в состоянии полного анабиоза, когда живая система является закрытой, она способна лишь испытывать элиминирующее действие естественного отбора под влиянием определенных агрессивных факторов среды. Однако направление этой элиминации всецело обусловлено развитием организма в метаболически активных фазах онтогенеза. Иначе говоря, процесс эволюции как таковой всегда ограничен периодами метаболически активной жизни, т. е. свойствен не всем, а только метаболически активным живым системам.
На основе общего определения живой системы открытая и закрытая живые системы могут быть определены следующим образом.
Открытая живая система — метаболически активная живая система, существующая на основе постоянного, активно осуществляемого обмена веществом и энергией с окружающей ее внешней средой, в ходе которого, в соответствии с программой развития системы, обеспечиваются ее сохранение и самовоспроизведение и создаются предпосылки к ее адаптивной эволюции.
Закрытая живая система — метаболически неактивная живая система, не осуществляющая обмена веществом с окружающей ее внешней средой и существующая в форме комплекса нефункционирующих жизнеспособных структур.
5. Периодический закон развития живых систем
В ходе биологической эволюции уровень функционально-структурной сложности живых систем изменяется двояким путем:
1) на дискретной основе, благодаря периодической агрегации доорганизменных или организменных систем в системы более высокого уровня структурной сложности. При этом структурная сложность систем увеличивается сразу на порядок: исходные системы в результате акта агрегации становятся подсистемами новой системы более высокого уровня структурной сложности. Именно таким путем происходило становление монобионтов, метабионтов и ценометабионтов;
2) путем медленных, постепенных изменений тех или иных элементов системы, в ходе которых новое качество накапливается в рамках определенного уровня структурной агрегации — монобионтного, метабионтного» или ценометабионтного. Так происходит адаптивная эволюция монобион-тов, метабионтов и ценометабионтов.
Целостная картина развития земной жизни представляет собою сочетание обоих этих путей и рисуется как сложный мультициклический процесс, подчиненный некоторому периодическому закону и состоящий в смене повторяющихся циклов, каждый из которых отличается от предыдущего более высоким уровнем структурной агрегации живой системы. В соответствии с этим выделяются три эры развития жизни на нашей планете, качественно различные по агрегатной структуре организмов: 1) монобионтная, в течение которой живые существа были представлены только монобионтами; 2) монометабионтная, в течение которой организмы были представлены монобионтами и метабионтами; 3) моно-мета-ценоме-табионтная, характеризующаяся присутствием в земной биоте одновременно всех трех структурно различных групп организмов — монобионтов, метабионтов и ценометабионтов — и продолжающаяся до настоящего времени.
Таким образом, в историческом развитии жизни на Земле происходили определенные периодические процессы, связанные с закономерной, циклической структурной агрегацией живых систем, составлявшей основу их адаптивного функционально-структурного усложнения. Эти периодические процессы могут быть описаны в виде периодического закона развития живых систем, сущность которого можно сформулировать следующим образом: эволюция организмов происходит на основе их периодической структурной агрегации, в ходе которой исходные доорганизменные или организменные системы становятся подсистемами новой организменной системы, более высокого структурного уровня.
Периодический закон развития живых систем отражает, следовательно, одну из наиболее общих особенностей их исторического развития — неравномерность и цикличность их эволюции.
В связи с задачами настоящего исследования — изучением экоморф» отражающих конкретную специфику организмов,— заметим, что организменный уровень организации живой системы в структурном отношении не является, следовательно, однозначным, а представлен тремя структурно различными уровнями, что имеет существенное значение при оценке особенностей экоморф самых различных организмов.
6. О техногенных вариантах живых систем
Искусственно созданный человеком, т.е. техногенный вариант живой системы, удовлетворяющий всем требованиям в ее общем определении, действительно был бы «живым» в том же смысле и в той же мере, как и естественные, природные живые системы. Не вдаваясь в технические детали этой проблемы и не касаясь ее экономических и социальных аспектов, отметим лишь ее общетеоретическую правомочность: искусственные, техногенные живые системы столь же возможны, как и любые созданные человеком техногенные объекты. Поскольку в случае создания техногенных живых систем речь идет фактически о моделировании свойственных природным живым системам конкретных функциональных особенностей, проблема техногенной жизни находится в рамках бионики и ее можно рассматривать как одну из технических задач, закономерна вытекающих из потребностей развивающейся человеческой цивилизации.
Выводы
Жизнь как, явление природы существует в форме дискретных живых систем. Основу дискретности живых систем составляет неделимая единица жизни — организм.
Живые системы обнаруживают значительное разнообразие, источником которого являются: 1) общий характер связей живой системы с внешней средой, 2) уровень функциональной организации системы, 3) уровень ее структурной агрегации и 4) способ организации процессов метаболизма. В связи с этим существенно деление живых систем соответственно на открытые и закрытые; организменные и надорганизменные; монобионтные, метабионтные и ценометабионтные; автобионтные и анавтобионтные. Эти деления неадекватны и не соподчинены друг другу; множественность их отражает многогранность процесса эволюции живых систем.
Разнообразие живых систем уменьшает число свойственных им общих особенностей, сводя его к трем взаимосвязанным главным свойствам живого: 1) наличию собственной программы развития живой системы, развертываемой на основе активно регулируемых информационных взаимосвязей ее с внешней средой, 2) иерархичности ее функционально-структурной организации и 3) ее высокой функционально-структурной сложности, особенно на молекулярном уровне.
Понятие живой системы относится к объектам весьма различного уровня сложности и может быть описано лишь в терминах биокибернетики а в соответствии с идеей системной организации. Живая система — иерархически организованная сложная система, имеющая собственную программу развития.
Эволюция организмов происходит на основе их периодической структурной агрегации, в ходе которой исходные доорганизменные или организменные системы становятся подсистемами новой организменной системы более высокого структурного уровня. Этот периодический закон развития живых систем отражает одну из наиболее общих особенностей их исторического развития — неравномерность и цикличность их эволюции.
Техногенные объекты, соответствующие определению живой системы, если таковые будут созданы, будут «живыми» в том же смысле и в той же мере, как и естественные, природные живые системы.
Антиэнтропичность живых систем непосредственно связана именно с их упорядоченностью и проявляется в том, что свойственный им уровень энтропии ниже возможного максимума, соответствующего термодинамически равновесному состоянию системы. Поддержание такого состояния, когда уровень энтропии системы отличается от максимума, возможно в двух случаях:
при положительном энергобалансе системы, когда потребляемая ею из окружающей среды энергия больше выделяемой, что, в частности, соответствует метаболически активному состоянию живой системы; в плане общей теории систем это состояние должно быть определено как состояние открытой системы;
при отсутствии такого положительного энергобаланса, если система способна сохранять определенный, отличный от максимума, уровень энтропии только за счет своих структурных антиэнтропических свойств, препятствующих увеличению энтропии при отсутствии активно осуществляемого организмом материального обмена с окружающей средой, что, в частности, соответствует метаболически неактивному состоянию живой системы, т. е. состоянию полного анабиоза; с позиций общей теории систем такое состояние отвечает состоянию закрытой системы.
Оба эти состояния, в частности структурные антиэнтропические свойства закрытых живых систем. Важно подчеркнуть, что антиэнтропичность не есть какое-то самостоятельное свойство живых систем; она представляет собою лишь следствие реально существующей высокой упорядоченности этих систем.
4. Высокая функционально-структурная сложность
Общим свойством всех живых систем, независимо от общего уровня их организации, является их высокая функционально-структурная сложность.
При сколь угодно простой макроорганизации живые системы неизменно остаются высокосложными за счет своей специфической микроорганизации: при любой сложности макроорганизации живой системы ее интегральная сложность всегда определяется ее микроорганизацией на молекулярном и субмолекулярном уровнях. Принимая во внимание все информационные, энергетические и материальные аспекты молекулярной и субмолекулярной организации живых систем, следует признать, что преобладание этой молекулярной составляющей в интегральной сложности живой системы в любом случае столь значительно, что совокупность всех ее остальных, т.е. надмолекулярных, компонентов, бесспорно, имеет лишь подчиненное значение. В этом смысле можно сказать, что по степени суммарной сложности шимпанзе меньше отличается от вируса табачной мозаики, чем молекулярная организация шимпанзе от ее надмолекулярной организации.
Основанием для такого заключения служат общеизвестные истины молекулярной биологии. Представление о степени информационной и функционально-структурной сложности живых систем на молекулярном уровне дают хотя бы общеизвестные особенности генетического кода организмов. Достаточно напомнить, например, что один ген, обеспечивающий кодирование только одного полипептида, содержит примерно 1,5 X. X 103 нуклеотидных пар в ДНК, а палиндромные последовательности в ДНК эукариот, представляющие собою области узнавания для рестриктаз, содержат до 1,2 • 103 нуклеотидных пар. В свете подобных цифр, число которых можно было бы увеличить, высокая информационная и функционально-структурная сложность молекулярной организации живых систем представляется достаточно очевидной.
Естественно, что пример с шимпанзе не претендует на абсолютную точность; однако он, несомненно, верно отражает нашу общую мысль о решающей роли молекулярных особенностей в определении общего интегрального уровня сложности живой системы. Это, разумеется не дает нам права сбрасывать со счетов элементы сложности надмолекулярных уровней; суть дела состоит лишь в том, что совокупность этих надмолекулярных элементов по своему относительному значению в суммарном выражении сложности живой системы любого ранга явно уступает молекулярной составляющей.
Такое положение вполне соответствует тому факту, что все происходящие в организмах важнейшие жизненные процессы, связанные с изменениями информационных, энергетических и материальных состояний живой системы, структурно обеспечены на молекулярном и субмолекулярном уровнях. Именно это обстоятельство делает возможным, в частности, существование неклеточных живых систем, т. е. вирусов, у которых вся функционально-структурная организация ограничена уровнем макромолекул и сравнительно несложных надмолекулярных агрегатов. В этом одно из важнейших свидетельств особой весомости молекулярного уровня в многоуровневой целостной конструкции живых систем любой сложности.
Высокая сложность живых систем, основанная на их упорядоченной, строго иерархической функционально-структурной организации, в сочетании с программным управлением всеми протекающими в рамках системы процессами информационного, энергетического и материального плана, и создает ту глубокую специфику живых систем, которая поднимает их на более высокий, качественно новый уровень организации, принципиально отличающий их от объектов неживой природы.
Принимая во внимание все сказанное, мы можем заметить, что с принципиальной стороны жизнь как общее явление во Вселенной представляет собою феномен скорее кибернетического, чем субстратного плана. В конечном итоге не белки и не нуклеиновые кислоты как конкретные полимерные субстраты, а прежде всего их свойства как накопителей и преобразователей информации явились необходимой основой формирования антиэнтропических самоповторяющихся и эволюционирующих, т. е. живых, систем: при наличии этих информационных свойств на какой-либо иной субстратной основе явление жизни в принципе могло бы развиться с таким же успехом.
Понятие живой системы относится к объектам весьма различного уровня сложности. Содержание этого понятия может быть определено лишь в терминах биокибернетики и в соответствии с идеей системной организации, на основе учета некоторых общих принципов функционирования живых систем, в частности в аспектах связей, контроля и управления. В соответствии с этим в общем определении живой системы не могут найти отражения конкретно-биологические особенности, характеризующие специфику тех или иных частных систем любого уровня: такие особенности могут быть учтены лишь в определениях живых систем конкретного уровня сложности, например в определении открытой и закрытой живых систем.
Принимая во внимание большое разнообразие уровней организации известных в настоящее время живых систем и существующие между ними различия в формах их связей с окружающей их средой, общее определение живой системы можно сформулировать в следующем виде.
Живая система — иерархически организованная сложная система, имеющая собственную программу развития, реализация которой обеспечивает сохранение системы на основе поддержания в ней определенного, отличного, от максимума уровня энтропии. Это определение подчеркивает, таким образом, все три основные особенности живой системы.
Биологи, несомненно, обратили внимание на то, что в приведенном выше определении живой системы отсутствуют указания на традиционно выделяемые свойства живого. Это объясняется тем, что эти «традиционные» свойства в большинстве своем могут относиться не ко всем, а только к открытым живым системам, с которыми их и было принято связывать. Иначе говоря, прежние определения живого, содержащие эти «традиционные» элементы, оставляли за пределами живого закрытые живые системы, соответствующие состоянию полного анабиоза. Естественно, что наше общее определение, относящееся ко всем без исключения живым системам, не может содержать в себе элементы, приемлемые только для открытых систем.
Как видно из приведенного определения, оно не содержит указания на нуклеиново-белковый характер субстратов живых систем. Это обусловлено тем фактом, что все три свойственные живым системам особенности принципиально могут быть реализованы не только на основе нуклеиново-белковых субстратов, но и на иной химической основе, в связи с чем этот нуклеиново-белковый вариант организации живой системы не может найти отражения в ее общем определении.
Попытки обосновать общее определение живой системы исходя из нуклеиново-белковой природы земной жизни закономерно приводят к ограничительному толкованию этого понятия. Одним из примеров такого подхода может служить хотя бы следующее определение: «Живыми называются такие системы, которые обладают нуклеиновыми кислотами и белками и способны сами синтезировать эти вещества. Это определение неприложимо к древнейшим ступеням возникновения жизни, а также к существующим, возможно, внеземным живым системам, которые могут быть устроены иначе». Как видно из этого текста, подобное определение требует немедленных оговорок по поводу ограниченности сферы его применения. Кроме уже содержащихся в этом тексте ограничений, авторам вышеприведенного определения приходится считать, что вирусы — это объекты, «состоящие из нуклеиновых кислот и белков, но не являющиеся живыми», что противоречит их же собственному определению живой системы. Таким образом, попытки определения живой системы на основе констатации нуклеиново-белковой природы земных организмов исходят из предпосылок частного характера, т.е. страдают геоцентризмом и в этом смысле отражают пройденный этап развития биологии.
Наше определение не содержит также и упоминания р метаболической деятельности живых систем, поскольку она не свойственна закрытым живым системам.
Далее, в нашем определении ничего не сказано о способности живых систем к самовоспроизведению. Это связано с тем, что не все живые системы являются самовоспроизводящимися. Достаточно вспомнить многие бесплодные касты насекомых, например рабочих пчел и термитов и т.п.: все они представляют собою примеры несамовоспроизводящихся живых систем. То же можно сказать и о многих живых системах надорганизмеииого уровня, к которым понятие «самовоспроизведения» неприменимо, каковы, в частности, временные, миграционные стаи многих животных — различных рыб, птиц и др.
Наше определение не содержит и указаний на способность живых систем к эволюции, поскольку эволюционировать способны не все, а только метаболически активные самовоспроизводящиеся живые системы. Что же касается закрытых живых систем, представленных организмами, находящимися в состоянии полного анабиоза, то они не способны ни к самовоспроизводству, ни к эволюции, поскольку существуют в форме нефункционирующей структурной системы организма. Эволюционное развитие адаптации, обеспечивающих биологическую допустимость анабиотических состояний, происходит лишь за счет тех или иных генетических изменений, возникающих в метаболически активных фазах онтогенеза, тогда как в состоянии полного анабиоза, когда живая система является закрытой, она способна лишь испытывать элиминирующее действие естественного отбора под влиянием определенных агрессивных факторов среды. Однако направление этой элиминации всецело обусловлено развитием организма в метаболически активных фазах онтогенеза. Иначе говоря, процесс эволюции как таковой всегда ограничен периодами метаболически активной жизни, т. е. свойствен не всем, а только метаболически активным живым системам.
На основе общего определения живой системы открытая и закрытая живые системы могут быть определены следующим образом.
Открытая живая система — метаболически активная живая система, существующая на основе постоянного, активно осуществляемого обмена веществом и энергией с окружающей ее внешней средой, в ходе которого, в соответствии с программой развития системы, обеспечиваются ее сохранение и самовоспроизведение и создаются предпосылки к ее адаптивной эволюции.
Закрытая живая система — метаболически неактивная живая система, не осуществляющая обмена веществом с окружающей ее внешней средой и существующая в форме комплекса нефункционирующих жизнеспособных структур.
5. Периодический закон развития живых систем
В ходе биологической эволюции уровень функционально-структурной сложности живых систем изменяется двояким путем:
1) на дискретной основе, благодаря периодической агрегации доорганизменных или организменных систем в системы более высокого уровня структурной сложности. При этом структурная сложность систем увеличивается сразу на порядок: исходные системы в результате акта агрегации становятся подсистемами новой системы более высокого уровня структурной сложности. Именно таким путем происходило становление монобионтов, метабионтов и ценометабионтов;
2) путем медленных, постепенных изменений тех или иных элементов системы, в ходе которых новое качество накапливается в рамках определенного уровня структурной агрегации — монобионтного, метабионтного» или ценометабионтного. Так происходит адаптивная эволюция монобион-тов, метабионтов и ценометабионтов.
Целостная картина развития земной жизни представляет собою сочетание обоих этих путей и рисуется как сложный мультициклический процесс, подчиненный некоторому периодическому закону и состоящий в смене повторяющихся циклов, каждый из которых отличается от предыдущего более высоким уровнем структурной агрегации живой системы. В соответствии с этим выделяются три эры развития жизни на нашей планете, качественно различные по агрегатной структуре организмов: 1) монобионтная, в течение которой живые существа были представлены только монобионтами; 2) монометабионтная, в течение которой организмы были представлены монобионтами и метабионтами; 3) моно-мета-ценоме-табионтная, характеризующаяся присутствием в земной биоте одновременно всех трех структурно различных групп организмов — монобионтов, метабионтов и ценометабионтов — и продолжающаяся до настоящего времени.
Таким образом, в историческом развитии жизни на Земле происходили определенные периодические процессы, связанные с закономерной, циклической структурной агрегацией живых систем, составлявшей основу их адаптивного функционально-структурного усложнения. Эти периодические процессы могут быть описаны в виде периодического закона развития живых систем, сущность которого можно сформулировать следующим образом: эволюция организмов происходит на основе их периодической структурной агрегации, в ходе которой исходные доорганизменные или организменные системы становятся подсистемами новой организменной системы, более высокого структурного уровня.
Периодический закон развития живых систем отражает, следовательно, одну из наиболее общих особенностей их исторического развития — неравномерность и цикличность их эволюции.
В связи с задачами настоящего исследования — изучением экоморф» отражающих конкретную специфику организмов,— заметим, что организменный уровень организации живой системы в структурном отношении не является, следовательно, однозначным, а представлен тремя структурно различными уровнями, что имеет существенное значение при оценке особенностей экоморф самых различных организмов.
6. О техногенных вариантах живых систем
Искусственно созданный человеком, т.е. техногенный вариант живой системы, удовлетворяющий всем требованиям в ее общем определении, действительно был бы «живым» в том же смысле и в той же мере, как и естественные, природные живые системы. Не вдаваясь в технические детали этой проблемы и не касаясь ее экономических и социальных аспектов, отметим лишь ее общетеоретическую правомочность: искусственные, техногенные живые системы столь же возможны, как и любые созданные человеком техногенные объекты. Поскольку в случае создания техногенных живых систем речь идет фактически о моделировании свойственных природным живым системам конкретных функциональных особенностей, проблема техногенной жизни находится в рамках бионики и ее можно рассматривать как одну из технических задач, закономерна вытекающих из потребностей развивающейся человеческой цивилизации.
Выводы
Жизнь как, явление природы существует в форме дискретных живых систем. Основу дискретности живых систем составляет неделимая единица жизни — организм.
Живые системы обнаруживают значительное разнообразие, источником которого являются: 1) общий характер связей живой системы с внешней средой, 2) уровень функциональной организации системы, 3) уровень ее структурной агрегации и 4) способ организации процессов метаболизма. В связи с этим существенно деление живых систем соответственно на открытые и закрытые; организменные и надорганизменные; монобионтные, метабионтные и ценометабионтные; автобионтные и анавтобионтные. Эти деления неадекватны и не соподчинены друг другу; множественность их отражает многогранность процесса эволюции живых систем.
Разнообразие живых систем уменьшает число свойственных им общих особенностей, сводя его к трем взаимосвязанным главным свойствам живого: 1) наличию собственной программы развития живой системы, развертываемой на основе активно регулируемых информационных взаимосвязей ее с внешней средой, 2) иерархичности ее функционально-структурной организации и 3) ее высокой функционально-структурной сложности, особенно на молекулярном уровне.
Понятие живой системы относится к объектам весьма различного уровня сложности и может быть описано лишь в терминах биокибернетики а в соответствии с идеей системной организации. Живая система — иерархически организованная сложная система, имеющая собственную программу развития.
Эволюция организмов происходит на основе их периодической структурной агрегации, в ходе которой исходные доорганизменные или организменные системы становятся подсистемами новой организменной системы более высокого структурного уровня. Этот периодический закон развития живых систем отражает одну из наиболее общих особенностей их исторического развития — неравномерность и цикличность их эволюции.
Техногенные объекты, соответствующие определению живой системы, если таковые будут созданы, будут «живыми» в том же смысле и в той же мере, как и естественные, природные живые системы.