Реферат на тему Принципы эволюции
Работа добавлена на сайт bukvasha.net: 2015-06-25Поможем написать учебную работу
Если у вас возникли сложности с курсовой, контрольной, дипломной, рефератом, отчетом по практике, научно-исследовательской и любой другой работой - мы готовы помочь.

Предоплата всего

Подписываем
Содержание.
ПРЕДИСЛОВИЕ РЕДАКТОРА ПЕРЕВОДА.
ПРЕДИСЛОВИЕ.
ГЛАВА 1. ИСТОРИЯ ФОРМИРОВАНИЯ ЭВОЛЮЦИОННЫХ ПРИНЦИПОВ.
1.1. Введение.
1.2. Постоянство, предначертанность и креационизм.
1.3. Запрограммированная эволюция.
1.4. Менее жестко запрограммированная эволюция.
1.5. Развитие дарвинизма и временный отход от него.
1.6. Выводы.
1.7. Рекомендуемая литература.
Глава 2. МЕХАНИЗМЫ НАСЛЕДСТВЕННОСТИ.
2.1. Механизм наследственности по Менделю.
2.2. Хромосомные и химические основы наследственности.
2.3. Неодарвинизм и популяционная генетика.
2.4. Выводы и резюме.
2.5. Рекомендуемая литература.
Глава 3. АДАПТАЦИЯ.
3.1. Введение.
3.2. Методология.
3.3. Меланизм у бабочек — подход a posteriori.
3.4. Примеры априорного подхода.
3.5. Изменчивость в природных популяциях; ее значение для адаптации.
3.6. Адаптации на благо группы и адаптации групп.
3.7. Адаптации самого полового размножения как такового и порожденные им адаптации.
3.8. «Эгоистичная» ДНК и гены в организмах.
3.9. Рекомендуемая литература.
Глава 4. ЭВОЛЮЦИЯ И РАЗВИТИЕ.
4.1. Введение.
4.2. Амплификация в процессе развития.
4.3. Канализация развития.
4.4. Заключения.
4.5. Рекомендуемая литература.
Глава 5. МАКРОЭВОЛЮЦИЯ.
5.1. Введение.
5.2. Таксономические категории и система классификации.
5.3. Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции.
5.4. Альтернативная точка зрения — прерывистая эволюция.
5.5. Сравнение теорий постепенной и прерывистой эволюции.
5.6. Проверка двух теорий эволюции.
5.7. Была ли эволюция человека постепенной или прерывистой?
5.8. Заключения.
5.9. Рекомендуемая литература.
ЛИТЕРАТУРА.
ПРЕДИСЛОВИЕ РЕДАКТОРА ПЕРЕВОДА.
Небольшая, но содержательная книга П. Кейлоу относится к серии книг, по которым изучают биологию на высшем, «третичном», уровне. В первую очередь она предназначена для студентов старших курсов и аспирантов, но немалую пользу может принести и лицам, самостоятельно продолжающим свое образование, да и специалистам, в том числе работающим в этой области.
Достоинство книг этой серии — современность. Авторы (и Кейлоу в том числе) смело доводят до аудитории новые, порой еще неустоявшиеся, а то и дискуссионные концепции. Такой подход нельзя не приветствовать, однако в нем таится и некоторая опасность. Излагая находящуюся в процессе разработки гипотезу, трудно оставаться объективным. В результате читатель может получить довольно искаженное представление о путях развития современной науки. Огульное охаивание, идущее от консерватора, и восторженный энтузиазм неофита здесь в равной мере опасны.
Приходится признать, что автор книги, представляемой советскому читателю, в ряде случаев не избежал этой опасности и кое-где акценты расставлены, на взгляд редактора, довольно субъективно.
В первую очередь это относится к взаимоотношениям между концепцией ламаркизма о наследовании приобретенных признаков и рабочей гипотезой Дарвина о пангенезе. Суть учения Ламарка не в наследовании приобретенных признаков (ибо если признак не приобретен, то нечего и наследовать), а в постулировании изначально адаптивной, приспособительной изменчивости.
Это и только это отличает ламаркизм от дарвинизма. Ведь всё признаки, вновь появляющиеся у потомков, возникают в результате изменения генов в предшественниках гамет родителей. Чем это не наследование приобретенных признаков?
С этой точки зрения и следует рассматривать рабочую гипотезу пангенеза, выдвинутую Дарвином. По Дарвину, геммулы (гипотетические частицы, несущие информацию о наследственных свойствах) переносятся в половые железы родителей, там включаются в гаметы и через них оказывают действие на потомков, то есть существует канал передачи информации от соматических клеток к генеративным. Но здесь нет приспособительной изменчивости, а следовательно, нет ламаркизма.
Кейлоу полагает, что Стеель, допуская перенос вирусами генетической информации от соматических клеток родителей к гаметам, вновь обращается к ламаркизму. Однако это скорее возврат к пангенезу Дарвина. Если механизм передачи информации, предложенный Стеелем, будет обнаружен в экспериментах и окажется, что он имеет какое-то значение для эволюции, то это лишний раз подтвердит старую истину о том, что гений, даже ошибаясь, не бывает неправ на все 100%. Нелишне напомнить, что Дарвин сопоставлял свои гипотетические геммулы с возбудителем оспы (теперь-то мы знаем, что это вирус!).
В последнее время ведутся ожесточенные дискуссии по поводу так называемой теории нейтральности (венцом которой является гипотеза эгоистичной ДНК, selfish DNA), о роли в эволюции крупных изменений генома, приводящих к существенному изменению фенотипа (макромутации), и о гипотезе прерывистого равновесия, возвращающей нас к скачкообразному происхождению видов по Де Фризу. П. Кейлоу также уделяет этим построениям немало внимания.
Следует учесть, что при оценке той или иной концепции необходимо устанавливать, в какой степени эта концепция касается основного принципа теории Дарвина — возникновения адаптивных (приспособительных) признаков в результате отбора ненаправленных наследственных изменений. Уже современники Дарвина понимали, что этот принцип является для эволюции основным. Так, еще Гельмгольц, выступая в 1869 г. на съезде естествоиспытателей в Инсбруке, говорил: «В дарвиновой теории заключена существенно новая, творческая идея. Она показывает, что целесообразность в строении организма могла возникнуть без вмешательства разума, в силу слепого действия одних законов природы».
В то же время Дарвин знал и писал об изменениях признаков, не снижающих жизнеспособность и фертильность организма и не подпадающих под действие отбора (нейтральных, как мы их сейчас называем). Оценка их роли в эволюционных изменениях— предмет дискуссии. Но даже если мы вслед за сторонниками теории нейтральности во главе с Кимурой примем, что нейтральные мутации — основной источник разнообразия в природе, то дарвиновский принцип эволюции сохранится. Целесообразность без отбора нельзя объяснить, не привлекая «вмешательства разума». Поэтому нейтральность и селекционизм не противоречат друг другу, они попросту лежат в разных плоскостях. Говорить о том, что теория нейтральности вытесняет или хотя бы дополняет дарвинизм, — заблуждение. С равным успехом можно пытаться заменить в диете один витамин другим (например, аскорбиновую кислоту сс-токоферолом).
Второе яблоко раздора современных эволюционистов — роль макромутаций в эволюции. Действительно ли Дарвин признавал только мелкие, незначительные изменения? Быть может, правы Г. Де Фриз и Р. Гольдшмидт, постулирующие необходимость макромутаций, резко, скачком перекраивающих фенотип? Участники дискуссии нередко забывают, что именно после того, как были открыты мутации, этот вопрос в значительной мере потерял остроту. Всякое наследственное изменение скачкообразно— но в какой степени? У дрозофилы описано множество мутаций, в той или иной мере детерминирующих развитие крылового аппарата. Так, мутация cut приводит к возникновению вырезок на крыльях, vestigial — к их недоразвитию, apterous — к полной редукции. Где проходит граница между микро- и макромутацией? Да и существует ли она в природе вообще?
Примеры макромутаций, приводимые Гольдшмидтом, явно неудачны. Например, у гомозигот tetraptera вместо жужжалец развивается вторая пара крыльев. Но это не возникновение нового признака, а возврат к старому — возникновение атавизма, — ведь жужжальца представляют собой производное второй пары крыльев. Закрепление атавизма в эволюции — вещь довольно редкая и обычно не приводит к особым новшествам. Мы знаем, что волосы млекопитающих развились в филогенезе из роговых чешуи рептильных предков. Известны и случаи возврата к прежнему состоянию (чешуйки на хвостах крыс и мышей, роговые чешуи панголинов). Вопрос, по-видимому, корректнее поставить так: какова относительная роль крупных изменений фенотипа в эволюции? Дарвин считал эту роль не столь значительной, так как крупные изменения возникают реже мелких, а адаптивными бывают в еще более редких случаях. Приходится признать, что его оппоненты веских доводов contra до сих пор не представили и оперируют не фактами, а аналогиями и логическими рассуждениями. И положение о том, что эволюционная ценность мутации обратно пропорциональна степени ее феноти-пического проявления, по-прежнему остается в силе.
В еще большей мере это относится к модной ныне гипотезе прерывистого равновесия, согласно которой видообразование — скачкообразный процесс, идущий за счет макромутаций, а целесообразность возникает в результате отбора видов (межвидовая борьба вместо внутривидовой). Автор явно симпатизирует этой концепции, но и он вынужден признать, что сторонники ее пока оперируют лишь гипотетическими моделями (шар, катящийся по профилированной плоскости), аналогиями с онтогенезом (смена креода, резкое изменение нормы реакции), да привлекают еще давний довод о редкости или отсутствии промежуточных форм в палеонтологической летописи. Но аналогии с онтогенезом неправомерны: еще И. И. Шмальгаузен (Проблемы дарвинизма, Л., 1969, с. 404) справедливо указывал, что «в основе обеих форм развития лежат совершенно различные факторы». Онтогенез — процесс, детерминированный генетической программой развития, на протяжении которого факторы внешней среды могут включать тот или иной вариант этой программы. Здесь нет стохастического детерминизма, характерного для филогенеза.
Неполнота палеонтологической летописи во времена Дарвина теперь компенсируется небывалым количеством полученного материала. Дж. Г. Симпсон справедливо указывает, что постепенность видообразования — твердо установленный палеонтологией факт, независимо от того, что она наблюдается не во всех случаях. Это в особенности относится к антропогенезу. Сейчас уже обнаружены промежуточные формы между современным человеком и неандертальцем (Схул и Табун), неандертальцем и питекантропом, сами питекантропы, начиная от слоя Джетис — наглядный пример «очеловечивания». Кустистая, а не линейная филогения гоминид — довод против ортогенеза, а не дарвинизма.
Вся история с гипотезой прерывистого равновесия — еще один факт, свидетельствующий о том, что противники Дарвина плохо его знают и часто дискутируют не с дарвинизмом, а со своим превратным представлением о нем. В самом деле, Дарвин знал и о нейтральных признаках, и о крупных наследственных изменениях, но не абсолютизировал их. Ошибкой будет считать и то, что, согласно Дарвину и синтетической теории эволюции, видообразование идет непрерывно и постепенно. Достаточно напомнить слова самого Дарвина: «...я полагаю, что виды обладают довольно хорошо определенными границами, и ни в какой период не бывает неразрешимого хаоса изменяющихся и промежуточных звеньев...» («Происхождение видов», гл. VI). Иными словами, Дарвин понимал, что в процессе видообразования популяция проходит некую критическую точку — переход из одного вида в другой, который не может длиться долго, а потому имеет мало шансов оказаться зафиксированным в палеонтологической летописи. Сторонники гипотезы прерывистого равновесия утрировали это положение, доведя его до абсурда и противопоставив синтетической теории эволюции. По-видимому, лучший способ дискредитировать какое-либо новое направление — это придать ему чересчур большое значение, переоценить и противопоставить другим. Я полагаю, что теория эволюции впитает все новое и ценное из гипотезы прерывистого равновесия, отвергнув ее непомерные притязания, как это случилось в свое время с теорией Де Фриза. История повторяется.
Полагаю, что книга П. Кейлоу будет иметь успех у советского читателя уже только потому, что она дает представление об обстановке на переднем крае развития эволюционной теории. Кроме того, она содержит сведения о дискуссионных взглядах и проблемах, которые можно почерпнуть лишь из статей в периодических изданиях, доступных далеко не каждому.
Б. М. Медников
ПРЕДИСЛОВИЕ.
За последние несколько лет появился ряд новых книг по эволюционной биологии. Однако они по большей части либо очень велики, либо слишком специальны. Настоящая книга — попытка создать общий очерк небольшого объема для студентов. Конечно, ограниченный объем требует тщательного отбора материала, поэтому мы стремились сосредоточить внимание на основных принципах, как теоретических, так и практических, не вдаваясь в излишние подробности. Иногда небольшой объем означает также, что автор рассчитывает на читателя, уже имеющего определенную подготовку, однако я надеюсь, что к данному случаю это не относится: я, безусловно, старался написать нечто в равной степени понятное и непосвященному, и хорошо осведомленному читателю. Что касается библиографии, то я по возможности ссылаюсь на обзоры, а не на оригинальные работы, и поэтому цитирование той или иной работы отнюдь не следует воспринимать как указание на приоритет ее автора.
При изложении материала я принял несколько вольный исторический подход не только потому, что читать книгу при этом становится интересней, но и потому, что рассматриваемый в ней предмет, равно как и читатель, к которому она обращена, развивались под критическим оком процесса отбора. Возникали проблемы, создавались гипотезы для их объяснения, подбирались фактические данные для проверки этих гипотез, обнаруживались новые проблемы, строились новые гипотезы и так далее. Я надеюсь, что это в какой-то мере ощущается при чтении книги и что в конечном итоге читатель получит ясное представление о том, что такое научный подход и как его применяют в эволюционной биологии.
П. К.
ГЛАВА 1. ИСТОРИЯ ФОРМИРОВАНИЯ ЭВОЛЮЦИОННЫХ ПРИНЦИПОВ
1.1. Введение.
Существуют два твердо установленных положения относительно живой природы: во-первых, в ней заключено богатейшее разнообразие животных, растений и микроорганизмов, а во-вторых, все живые организмы, по-видимому, хорошо приспособлены к тому, чтобы решать проблемы, встающие перед ними в той среде, в которой они обитают. Можно представить себе, что либо все признаки живых организмов постоянны и оставались таковыми с тех пор, как зародилась жизнь, либо эти признаки способны изменяться и их разнообразие и адаптивность прогрессивно развертывались с течением времени. Этот последний динамический процесс теперь называют эволюцией. Относительно механизмов эволюции высказывались различные предположения— от вмешательства богов до естественного отбора. Последняя точка зрения, впервые высказанная Дарвином, в настоящее время занимает господствующее положение, и цель этой книги состоит в том, чтобы суммировать ту сложную совокупность мыслей и фактов, которые оказались с ней связанными, особенно за последние сто лет. Однако для того чтобы сделать это должным образом, необходимо оценить исторические и философские предпосылки, на фоне которых Дарвин совершил переворот в науке, а поэтому в настоящей главе будет дан краткий исторический обзор и сделаны некоторые замечания философского характера.
1.2. Постоянство, предначертанность и креационизм.
Выражение «nullae speciae novae» было чрезвычайно ходовым у первых систематиков, в том числе у Линнея (1707— 1778). Считалось, что виды созданы творцом, как это описано в Книге Бытия, и поэтому они неизменны. Считалось также, что» они так хорошо приспособлены ко всем превратностям своей повседневной жизни потому, что, создавая их, творец предусмотрел соответствие каждого из них его особым функциям в приро-те. Джон Рей (Ray, 1627—1705)—священник, естествоиспытатель и один из первых систематиков — видел в приспособленности видов доказательство существования творца, а позднее Уильям Пэйли (William Paley, 1743—1805) придал этой «аргументации от плана строения» (Argument from Design) еще большую определенность. Животные и растения, утверждали они, — это удивительные механизмы, более удивительные, чем любая из машин, сконструированных человеком. (Позднее появилась более утонченная версия этой аргументации, в которой на первый план выдвигалась гармоничность формы, а не ее полезность. Эти две точки зрения соответственно получили названия идеалистической и утилитарной.)
Однако креационистская позиция породила ряд биологических и даже теологических и философских проблем. Так, например, было обнаружено много ископаемых остатков организмов, которые более не существуют на Земле, а между тем нельзя было представить себе, чтобы совершенные создания всемогущего творца могли когда-либо устареть. Чтобы выйти из этого затруднительного положения, Кювье (Cuvier, 1769—1832), чрезвычайно влиятельный французский биолог, выдвинул теорию катастроф. В результате ряда катаклизмов, одним из которых был всемирный потоп, некоторые из созданных творцом видов были уничтожены. Еще одно затруднение порождали признаки, казавшиеся бесполезными, поскольку опять-таки едва ли можно было допустить, что из рук сверхразумного создателя могло выйти нечто несовершенное. Другой французский биолог Бюффон (Buffon, 1707—1788) писал о свинье: «У нее имеются явно бесполезные части или, вернее, части, которые она не может использовать: в скелете конечностей все фаланги пальцев полностью сформированы, а между тем они ей совершенно не нужны».
Пытаясь объяснить существование таких бесполезных признаков, Бюффон высказал предположение, что Высшее существо создало совершенные типы, воплощенные в первоначальных видах, от которых впоследствии в результате процесса гибридизации и вырождения возникли новые виды. Так, предполагалось, что осел — это выродившаяся лошадь, а обезьяна — выродившийся человек. Но это означало отказ от допущения неизменности видов, и концепция nullae speciae novae исчезла из последнего пересмотренного издания линнеевского труда «Sys-tema naturae». («Аргументация от плана строения» грешит также более глубокими теологическими и философскими недостатками, относящимися к происхождению разного рода несовершенных признаков и к образу творца как некоего человекоподобного конструктора, но этих вопросов мы касаться не будем.)
1.3. Запрограммированная эволюция.
Даже в средние века некоторые ученые считали сотворение мира, описанное в Книге Бытия, мифом. Августин (353—430) уподоблял труд создателя прогрессивному росту дерева; подобным же образом Фома Аквинский (1225—1274) рассматривал сотворение мира как процесс постепенного развертывания возможностей, заложенных творцом в материю.
Именно в этом смысле Шарль Боннэ (Charles Bonnet, 1720— 1793) впервые использовал термин эволюции (e-volvo — развертывать), перенося представление о прогрессивном эмбриогенезе (у отдельных особей) на развитие видов. Боннэ и Тревира-нус (Treviranus, 1776—1853), прославившийся как создатель термина «биология», придерживались мнения, что план строения всех форм жизни содержался в виде зародышей в первом живом существе. Однако Боннэ представлял себе это развертывание как выполнение строго детерминированной программы изменений, тогда как Тревиранус рассматривал эволюцию как реализацию тех из бесчисленного множества возможных форм (запрограммированных в каждом организме творцом), которые будут обладать свойствами, необходимыми для того, чтобы выжить в изменяющемся мире. Таким образом, все ныне существующие и вымершие виды возникли в результате взаимодействия организма и среды.
Подобные взгляды на мир как на гигантский организм, наделенный неким таинственным импульсом к развитию, которые высказывал еще Платон (427—347 гг. до н. э.), можно найти в трудах таких эволюционистов XVIII и даже XIX веков, как Гете (1749—1831), Кант (1724—1804) и Чемберс (Chambers, 1802—1871) —так называемых натурфилософов.
1.4. Менее жестко запрограммированная эволюция.
Но поколенья множились, цвели, Усилились и члены обрели, Восстал растений мир, и средь обилья Разнообразной жизни в ход пошли Животных ноги, плавники и крылья.
Так писал дед Чарлза Дарвина — Эразм Дарвин (1731— 1807) в своей поэме «Храм природы» (Лондон, 1806). Э. Дарвин. Храм природы. Перевод Н. А. Холодковского. — М.: изд-во АН СССР, 1954, с. 21.
В этой и в другой своей книге (Zoonomia, London, 1794—1796) он предложил менее жесткую систему детерминированной эволюции, основанную на приобретении новых признаков самими организмами. Ламарк (Lamarck, 1744—1820) в своей «Философии зоологии» (Philosophic Zoologique, Paris, 1809) сформулировал очень сходную теорию. Чарлз Дарвин всегда подозревал, что Ламарк заимствовал идеи Эразма Дарвина, но это никогда не было твердо установлено.
Обе теории были основаны на трех главных принципах.
1. Считалось, что все организмы обладают врожденной способностью развиваться в сторону усложнения и совершенствования формы. Эразм Дарвин считал, что эта «способность совершенствоваться за счет своей собственной врожденной активности» дарована им богом; Ламарк же полагал, что «жизненная сила» проистекает из самой организации живых существ. Через организм протекают тонкие флюиды — тепловые и электрические,— не только поддерживающие, но и усиливающие его упорядоченность.
2. Все организмы имеют некую внутреннюю склонность (то, что Ламарк называл внутренним стремлением), которая заставляет их выполнять действия, достаточные для того чтобы удовлетворять требованиям (обеспечивать выживание), предъявляемым изменяющейся средой. Среда не вызывает изменений, она вызывает потребность в изменении, которую организм осознает и в соответствии с которой он действует. Эразм Дарвин выходил из положения, привлекая для объяснения этой внутренней склонности некое таинственное начало, а Ламарк — тонкие флюиды.
3. Признаки, приобретенные организмом в результате употребления того или иного органа в каких-то новых целях, передаются от одного поколения другому благодаря процессу, которому ни Дарвин, ни Ламарк не дали четкого определения. И напротив, признаки, утраченные вследствие неупотребления в одном поколении, не появляются в последующих поколениях.
Используя эти принципы, Ламарк объяснял эволюцию щупалец, имеющихся на голове улитки, следующим образом. Когда улитка ползет, она ощущает потребность прикасаться к находящимся перед ней предметам и прилагает усилия, чтобы дотронуться до них самой передней частью головы. При этом она посылает в выросты головы тонкие флюиды. В конце концов в эти выросты проникают нервные волокна и они превращаются в постоянные щупальца. Более того, приобретенные таким образом щупальца передаются от родителей потомкам, то есть они наследуются. Читателю, возможно, знакомо аналогичное объяснение эволюции шеи у жирафа, и он в состоянии представить себе, что подобным образом можно объяснить возникновение всех приспособлений. Следует, однако, указать, что наследуемость приобретенных признаков — лишь часть теории эволюции Ламарка. Другая, более важная, ее часть относится к тому, как эти признаки приобретаются, а именно — в соответствии с тем, что необходимо организмам для того чтобы выжить в данное время или даже в будущем, причем они ощущают эти потребности каким-то таинственным путем.
1.5. Развитие дарвинизма и временный отход от него.
Чарлз Дарвин (1809—1882) не критиковал Ламарка —этого «заслуженно знаменитого естествоиспытателя», как он называет его в историческом очерке к четвертому изданию «Происхождения видов» — за его идею об эволюции жизни или даже за его идею о том, что признаки, приобретенные организмом в течение его жизни, могут передаваться по наследству его потомкам. Вопрос, по которому Дарвин расходится с Ламарком и с собственным дедом, относится к механизму эволюции. Ламарк и Эразм Дарвин допускали вмешательство каких-то таинственных сил, наделяющих организмы способностью самим выбирать для себя именно те признаки, которые обусловили эволюционный прогресс в направлении все более и более совершенных форм адаптации. Но можно ли представить себе, чтобы организмы могли отличать полезные приобретенные признаки (которые обычно редки) от вредных (которые довольно обычны, как, например, раны, переломы конечностей, последствия заболеваний и т. п.) без вмешательства какой-то в высшей степени изощренной и разумной силы, способной устанавливать эти различия? И как можно говорить, что организмы стремятся к совершенству, не обращаясь вновь к предначертанному плану и к его создателю? Чарлз Дарвин хотел дать объяснение органической эволюции, не прибегая ни в явной, ни в скрытой форме к таким субъективными и антропоморфистским представлениям. В своей книге «Происхождение видов» (впервые изданной Джоном Мерреем в 1859 г.) Дарвин заменил идею о направленной изменчивости (вызываемой «внутренним стремлением») идеей случайной изменчивости, передаваемой по наследству. О происхождении этой изменчивости он высказывался несколько неопределенно, полагая, что она, возможно, обусловлена воздействиями среды на половые железы, и, как мы увидим ниже, столь же неопределенно он высказывался относительно механизма наследования изменчивости. Однако он совершенно ясно сформулировал, что каким бы образом ни возникала изменчивость, это происходило вне всякой зависимости от потребностей организма. Кроме того, Дарвин считал, что та изменчивость, которая лежит в основе эволюции, слагается из очень небольших непрерывных отклонений от первоначальной формы. О допускал возможность крупных внезапных изменений, но считал, что они редки и в большинстве случаев ведут к уродствам, По его мнению, этого, в сущности, и следовало ожидать, по скольку полагать, что какое-либо крупное изменение в морфолог гии или физиологии могло бы а) адаптироваться к уже сущест вующей организации того или иного организма и б) обладать селективным преимуществом, «значило бы перейти в область чудесного, покинув область Науки». Наконец, вместо ламар ковского таинственного стремления к совершенству Дарвин представлял эволюцию как весьма обычный процесс, лишенный всякой таинственности. Животноводы и садоводы, говорил он, путем отбора и разведения отклоняющихся индивидуумов получили огромное разнообразие форм домашних животных и ра тений. Это был искусственный отбор. Дарвин полагал также что борьба между организмами в перенаселенном мире за огра-' ничейные ресурсы неизбежно приводит к тому, что выживают только наиболее приспособленные и что это должно вести к сво-его рода естественному отбору вариантов, — мысль, на которую) его натолкнули труды Мальтуса по экономике (Malthus, A: Essay on the Principle of Population, London, 1798).
Таким образом, суть дарвиновской теории механизма эво люции сводилась к следующему: 1) у особей в пределах каждо го отдельного вида наблюдается значительная, но непрерывна изменчивость по морфологическим и физиологическим призна кам; 2) эта изменчивость возникает случайным образом и насле дуется; 3) популяции животных и растений обладают значительной способностью к увеличению; 4) однако необходимые им ресурсы ограничены, и поэтому особи данной популяции борются1 за собственное существование и за существование своих потомков; 5) поэтому только некоторые (те, кого Дарвин назвал наиболее приспособленными) выживают и оставляют потомков, обладающих теми же самыми признаками; 6) в результате такого естественного отбора наиболее приспособленных представители данного вида становятся все лучше и лучше адаптированными к окружающим условиям.
Из предпосылок, на которых Дарвин строил свои рассуждения,— изменчивости, наследственности и ограниченности ресурсов— следствие (эволюция путем естественного отбора): вытекает автоматически; кому-то может показаться — даже; слишком автоматически, чтобы все это можно было серьезно принять в качестве научной гипотезы. Выживают приспособленные, потому что по определению те, кто выживает, — приспособлены; если дарвинизм свести только к этому, то он представляется довольно поверхностным, а аргументация тавтологической. Между тем это не так. Во-первых, влияние тех или иных признаков на выживание и воспроизведение не постоянно и одинаково, а варьирует в зависимости от того, к какому таксону принадлежит данный организм, и от его местообитания; поэтому приспособительную ценность каждого отдельного признака необходимо исследовать и объяснить (см. гл. 3). Во-вторых, «выживание наиболее приспособленных» — всего лишь незрелое и непродуманное обобщение (заимствованное Дарвином у Герберта Спенсера, 1820—1903) того, что было задумано Дарвином. Это вершина теоретического «айсберга», под которой лежали и лежат менее очевидные допущения об источнике изменчивости и механизме наследственности (см. гл. 2). В принципе эти допущения могут оказаться ошибочными, и они поддаются проверке; именно это (то есть подверженность теории ошибкам и возможность ее проверки) делает дарвинизм интересным с научной точки зрения.
Но, признавая логическую строгость и научную приемлемость дарвинизма, можно ли считать эту теорию действительно правдоподобной? Можно ли ожидать, что отбор случайных изменений приведет к возникновению тех сложных и интегрированных механизмов, которые имеются у организма? Пэйли (Paley) заметил, что один вид такого органа, как глаз, вылечит от атеизма любого, ибо невозможно представить себе, чтобы такая сложная и высокоинтегрированная система могла возникнуть случайно. Сам Дарвин указывал, однако, что если в живой природе существует непрерывная последовательность от простых рудиментарных органов, воспринимающих свет, до глаза позвоночных и если каждый из таких органов приносит некоторую пользу своему носителю, то возражение Пэйли снимается, поскольку при этом нет необходимости считать, что глаз возник сразу; его развитие могло проходить через ряд последовательных ступеней, связанных между собой непрерывностью, создаваемой наследственностью, при содействии со стороны естественного отбора. Тем не менее в частных беседах Дарвин высказывался не столь уверенно, признавшись одному из своих коллег, что хотя одного лишь созерцания глаза было недостаточно, чтобы сделать его верующим, при виде глаза он не раз обливался холодным потом. Даже при наличии таких последовательных ступеней трудно поверить, что тот или иной тонко сбалансированный и превосходно скоординированный орган мог возникнуть в результате множества случайных событий, и многие до сих пор придерживаются точки зрения Пэйли. (Ридли (Ridley 1982) дал исторический обзор затруднений, с которыми сталкивается теория Дарвина, когда дело доходит до сложных коадаптированных органов.Ц
Теперь на нашей стороне по крайней мере фактор времени— чего не было у Дарвина. Иными словами, мы теперь можем с достаточной степенью уверенности утверждать, что с момента возникновения первого живого организма прошло примерно 3500 млн. лет, а с тех пор, как у животных, подобных плоским червям, появились первые рудиментарные фоторецепторы, прошло более 1000 млн. лет (табл. 1.1). За эти невообразимо длительные периоды времени даже невероятные события становятся мыслимыми. Однако Уильям Томсон (W. Thomson, 1824—, 1907), впоследствии лорд Кельвин (Kelvin), сократил время, которым мог оперировать Дарвин, берясь доказать, что возраст Земли не может превышать 100 млн. лет. Эта ошибочная оценка ввергла Дарвина и дарвинистов в состояние неуверенности и подорвала веру к ним, во всяком случае со стороны их современников.
Таблица 1.1. Принятая в настоящее время временная шкала для истории Земли и эволюции жизни.
| Эра | Период | Главные эволюционные события | Время2 | |
| 1 | Плейстоцен | Человек | 11.59 | |
| 10 | Плиоцен |Неоген | |||
| 30 | Кайнозой | Миоцен | ||
| 40 | Олигоцен | |||
| 60 | Эоцен > Палеоген | |||
| 75 | Палеоцен j | 11.00 | ||
| 135 | Мел | Млекопитающие | ——— | |
| 165 | Мезозой | Юра | Цветковые растения | |
| 205 | Триас | 9.30 | ||
| 230 | Пермь | Тайнобрачные | ||
| 250 280 | Пенсильвании Миссисипий | Карбон | Рептилии | ||
| 325 | Палеозой | Девон | Амфибии | 8.30 |
| Насекомые | ||||
| 360 | Силур | Наземные растения | 8.00 | |
| Рыбы | ||||
| 425 | Ордовик | Водоросли | ||
| 500 | Кембрий | Все главные типы | ||
| беспозвоночных | 6.00 | |||
| 3000 | Протерозой | Первые ископаемые | 1.00 | |
| Докембрий | (1000) | |||
| 10000 | Архей | Первое появление | 0.00 | |
| жизни | (полночь) | |||
| (3000—4000) | ||||
| Образование Земли | ||||
| (10000) | ||||
| 1 Время в миллионах лет от начала периода. | ||||
| г Время по 24-часовой шкале с момента возникновения жизни. | ||||
Сверх всего этого Дарвин связал свою теорию с механизмом наследственности, основанным на смешении наследственных задатков родительских особей, что было роковым для постулированных им мелких изменений. Он полагал, что вклады двух родительских особей смешиваются, давая потомство с промежуточными признаками. Однако на такой основе отбор благоприятных вариантов попросту невозможен, так как их в конечном счете неизбежно поглотили бы неблагоприятные варианты! Подумайте, что бы произошло, писал один из рецензентов «Происхождения видов» Флиминг Дженкин (Fleeming Jenkin, North Brit. Review, 1867, 149—171), с наследственными признаками белого человека, оказавшегося после кораблекрушения на острове, населенном неграми. Независимо от степени «превосходства», (слова рецензента) этих признаков с помощью простой арифметики можно показать, что всего за несколько поколений они были бы сведены на нет в результате смешения с признаками местного населения.
Для того чтобы справиться с затруднениями, возникшими в связи с представлениями о слитной наследственности, и с возражениями, выдвинутыми лордом Кельвином, Дарвину пришлось отвести в своей теории эволюции более важную роль наследованию признаков, приобретенных организмом в течение жизни. Таким путем вещество наследственности могло бы за короткое время ассимилировать относительно крупные и интегрированные изменения. Но при этом различие между дарвинизмом и ламаркизмом становится менее четким и вновь возникает старая проблема о том, почему одни приобретенные признаки оказываются закодированными в наследственном материале, а другие — нет. Из-за этого вера в дарвинизм еще больше ослабла, и в начале нашего века Норденшельд в своей классической «Истории биологии» (Nordenskiold, History of Biology, Tudor Publ. Co., 1929) посвятил большую главу упадку и даже полной отставке дарвинизма. «Дарвиновская теория происхождения видов, — писал он, — давно отброшена». Не говоря уже о переоценке длительности времени, которым располагала эволюция, необходимо было лучше представлять себе механизм наследственности, который мы рассмотрим в гл. 2 под рубрикой неодарвинизма (разд. 2.3). В течение короткого времени существовал также лагерь неоламаркистов (но см. разд. 2.2.4), отстаивавших естественный отбор вместе с приобретенными признаками. Например, Сент-Джордж Джексон Майварт в своей книге «К вопросу о происхождении видов» (St. George Jackson Mivart, «On the Genesis of Species», Macmillan, 1871) писал, что он не может представить себе, как путем постепенного накопления мелких изменений могла происходить эволюция, поскольку признаки в промежуточном состоянии бывают по большей части слабоадаптивными (см. разд. 5.3). Поэтому он высказал предположение, что эволюция происходит скачками и что основой для этих скачков служит приобретение полезных признаков. К этому вопросу мы еще вернемся в гл. 5.
1.6. Выводы.
В заключение следует еще раз подчеркнуть различие между а) точкой зрения о том, что жизнь возникла в процессе эволюции, и б) теориями о том, как происходила эволюция. К тому времени, когда было опубликовано «Происхождение видов», многие были готовы признать внушительный ряд данных, свидетельствующих в пользу эволюции, и в частности:
1) существование ископаемых остатков, свидетельствующих об изменениях фауны и флоры с течением времени;
2) сходство, лежащее в основе морфологии, физиологии и эмбриологии разных видов, наводящее на мысль об их общих предках;
3) географическая прерывистость в фаунах и флорах, связанная с физическими преградами (например, Австралия с ее своеобразными сумчатыми млекопитающими или острова — подобно Галапагосской группе, которые посетил и описал Дарвин,— с характерными для них одних видами растений и животных), заставляла предполагать естественный процесс изменения признаков у видов, оказавшихся «в ловушке» в этих изолированных областях.
Были возможны, однако, и многие другие объяснения эволюционного изменения. Дарвин выдвинул одно объяснение — естественный отбор случайных изменений, — менее антропоморфное, чем другие, и подтверждавшееся рядом наблюдений, в частности:
1) у особей, составляющих природные популяции, наблюдается значительная и притом часто непрерывная изменчивость;
2) путем отбора такой изменчивости животноводы добиваются внушительных морфологических и физиологических изменений у домашних животных;
3) адаптации основаны на модификации обычных принципов и зачастую бывают не так хороши, как могли бы быть, если бы они создавались de novo, без ограничений, налагаемых пред существующими структурами. Несовершенные решения, явно бесполезные признаки и рудиментарные органы, служат — несколько парадоксальным образом — доводом в пользу процесса эволюции, основанного на естественном отборе случайных изменений в рамках ограничений, создаваемых предсуществующими формами. Их трудно совместить с концепцией всемогущего творца.
1.7. Рекомендуемая литература.
Общая история биологии хорошо изложена в книге Смита (9). Полные обзоры истории, философии и социального значения дарвинизма содержатся в книгах Джиллеспи (2), Моора (4) и Рьюза (7, 8). Сведения о Дарвине как человеке и его путешествии на «Бигле» можно найти у Мурхеда (5), Джорджа (11 и Ховарда (3).
Глава 2. МЕХАНИЗМЫ НАСЛЕДСТВЕННОСТИ.
2.1. Механизм наследственности по Менделю.
2.1.1. Основные принципы.
Дарвин сознавал, что для правильной оценки эволюционных процессов очень важно отчетливо понимать явление наследственности. А между тем он завяз в теории, согласно которой наследственность работала против естественного отбора, «разбавляя», а не «концентрируя» благоприятные варианты. Человеком, разработавшим теорию наследственности, более совместимую с дарвиновской эволюцией, и заложившим основы современной генетики, был Грегор Мендель (Gregor Mendel, 1822—1884) В 1865 г. он сообщил о результатах своих экспериментов по гибридизации растений на заседании Общества естествоиспытателей в Брюнне (Брно), а на следующий год опубликовал их в «Трудах» этого общества. Однако его работа почти не привлекала внимания до тех пор, пока в 1900 г. Корренс (Correns) в Германии, де Фриз (de Vries) в Голландии и Чермак (Tschermak) в Австрии не открыли обнаруженные Менделем закономерности вторично и не оценили их значение и важность для теории Дарвина.
Изучая горох (Pisum sativutn), Мендель сумел выбрать у этого растения ряд особенностей, удобных для проведения экспериментов: 1) оно представлено многочисленными формами, четко отличающимися друг от друга по признакам, относящимся к семенам и к растению в целом; 2) предоставленные самим себе, эти растения могут самоопыляться, так как в каждом цветке имеются и тычинки, и пестики; 3) возможно проведение контрольных скрещиваний путем удаления тычинок из цветков (для предотвращения самоопыления) и опыления цветков пыльцой,, взятой от других растений.
Мендель выбрал для своей работы несколько признаков (табл. 2.1) и путем многократных самоопылений вывел чистые линии по этим признакам. Таким образом он установил, что растения размножаются по этим признакам в чистоте на протяжении многих поколений, из чего следовало, что контролирующие эти признаки наследственные факторы, по-видимому, очень стабильны. Проводя дальнейшие экспериментальные скрещивания, Мендель заметил, что при скрещивании двух чистых линий, различающихся по какой-либо паре признаков, все потомки (первое дочернее поколение, или f1) несут только один из этих признаков. Если, однако, скрещивать между собой растения этого первого поколения, то следующее поколение (поколение F2) содержит растения с признаками своих родителей, а также растения с исходными признаками, отсутствовавшими в поколении fi. Это означает, что признаки исходных растений выщепляются в F2, а следовательно, в fi не происходит слияния генетического материала. Наконец, Мендель обнаружил, что в поколении F2 признаки одного и другого типа появляются в определенном соотношении — примерно 3:1.
Таблица 2.1. Признаки, изучавшиеся Менделем (доминантные признаки набраны прописными.
| Признак | Варианты |
| Форма семян | ГЛАДКИЕ и морщинистые |
| Окраска семян | ЖЕЛТЫЕ и зеленые |
| Оболочка семени | ОКРАШЕННАЯ и белая |
| Бобы Окраска боба | ГЛАДКИЕ и морщинистые ЗЕЛЕНАЯ и желтая |
| Цветки | ПАЗУШНЫЕ и верхушечные |
| Стебель | ДЛИННЫЙ и короткий |
Мендель интерпретировал эти данные следующим образом: 1) наследственные детерминанты (он называл их факторами, а теперь мы называем их генами) дискретны; 2) они не сливаются; 3) у каждой взрослой особи имеется по два таких детерминанта (теперь их называют аллелями) для каждого признака, и они распределяются между гаметами — по одному на каждую гамету; 4) каждая взрослая особь может нести либо два идентичных аллеля (такую особь теперь называют гомозиготной), либо два разных аллеля (гетерозиготная особь), и в последнем случае экспрессия одного аллеля доминирует над экспрессией Другого (табл. 2.1); 5) гаметы соединяются случайным образом. Пункт 3 часто называют первым законом Менделя. Он означает, что гетерозиготные родительские особи производят в равных количествах гаметы, содержащие контрастирующие аллели.
Мендель обозначал пары генов буквами; доминантные аллели— прописными, а рецессивные — строчными. На рис. 2.1 эти обозначения использованы для наглядного изображения результатов одного из его экспериментов и их истолкования. Мендель скрещивал растения гороха с гладкими (доминантный признак— R) и морщинистыми (рецессивный признак — г) семенами и получил в поколении F2 5474 гладких и 1850 морщинистых семян, то есть отношение гладких семян к морщинистым составляло 2,96: 1, что совпадает с предсказанным соотношением 3:1.
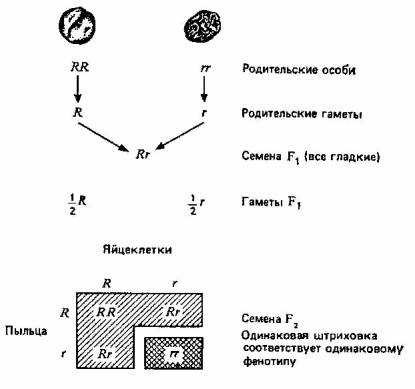
Предсказанное соотношение = 3 гладких :1 морщинистое
Рис. 2.1. Один из экспериментов Менделя по скрещиванию растений, различающихся по одной паре признаков.
Далее Мендель обнаружил, что если исходные растения, и с-! пользованные в скрещивании, различаются еще по двум контрастирующим признакам, то эти признаки в поколении fs также расщепляются, но при этом не обязательно остаются связанными с первой парой признаков. На основе этого Мендель сформулировал свой второй закон, или принцип независимого распределения: гены, детерминирующие разные признаки, распределяются по гаметам независимо друг от друга. (Последнее далеко не всегда справедливо, но это было установлено позднее.) Один из экспериментов Менделя по изучению наследования двух признаков и истолкование полученных данных представлены на рис. 2.2. Интересные результаты наблюдаются в поколении F2. Существует две возможности: 1) признаки, полученные от одной из родительских особей, передаются совместно, и в таком случае следует ожидать в поколении F2 семена только двух типов — гладкие-желтые и морщинистые-зеленые — в соотношении 3:1, как и прежде (рис. 2.1); 2) эти признаки передаются независимо, как показано на рис. 2.2. В последнем случае в fz должно быть 4 типа семян в соотношении 9 гладких-желтых (доминантный—доминантный), 3 гладких-зеленых (доминантный—рецессивный), 3 морщинистых-желтых (рецессивный—доминантный), 1 морщинистый-зеленый (рецессивный—рецессивный). Мендель получил семена этих четырех типов в соотношении 315 : 108 : 101 : 32, что хорошо соответствует пункту 2.
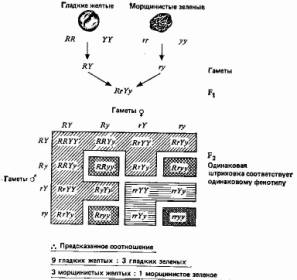
Рис. 2.2. Еще один эксперимент Менделя, в котором скрещивали растения, различающиеся по двум парам признаков.
2.1.2. Осложнения.
Как и можно было ожидать, дальнейшие исследования показали, что генетический механизм сложнее, чем представлял себе Мендель. Тем не менее ни одно из этих осложнений не опровергает основных допущений Менделя о дискретности генов и их большой стабильности. Ниже кратко изложены некоторые наиболее значительные осложнения.
1. Доминирование. Оперение кур андалузской породы отливает синим, но среди цыплят всегда попадаются черные особи и особи с белыми пятнами. На самом деле андалузки гетерозиготны по окраске оперения, но черная окраска не полностью доминирует над пятнистой. Это не может служить доводом в пользу слитной наследственности, потому что в потомстве от скрещиваний между гетерозиготными особями наблюдается выщепле-ние пятнистой и черной окраски в соответствии с менделевски-ми соотношениями.
Теперь мы знаем, что степень доминирования одного гена над другим зависит от его экспрессивности, а не от каких-либо существенных различий между генетическими системами. При кодоминантности оба аллеля экспрессируются в одинаковой мере (то есть аддитивны), именно это и происходит у кур андалузской породы. При частичном, или неполном, доминировании один из аллелей не активен (не экспрессируется), но другой активен полностью, так что в фенотипе эффект проявляется наполовину. При полном доминировании (то, что наблюдал Мендель) эффект одного аллеля эквивалентен эффекту двух аллелей, а при сверхдоминировании эффект одного аллеля выражен в большей степени, чем эффект двух таких аллелей в гомозигот-ном состоянии. Картина еще больше осложняется в тех случаях, когда экспрессия аллелей, находящихся в одной части (локусе) генного набора, регулируется экспрессией аллелей, находящихся в других локусах. Это явление известно под названием эпис-таза. Например, при скрещивании двух гомозиготных мышей, одна из которых доминантна по окраске шерсти — агути (серая шкурка), а другая — рецессивна (черная шкурка), все потомство должно было бы иметь окраску агути. Большая часть мышат действительно относится к типу агути, однако встречаются и альбиносы, причем частота их выше той, которую можно былом бы отнести за счет мутаций. Происходит это потому, что на ок-" раску шкурки влияет еще и другая пара генов, находящихся в другом локусе. Доминантный аллель в этом локусе определяет нормальную окраску, детерминированную аллелями главного локуса, а рецессивный аллель определяет альбинизм, и любая мышь, гомозиготная по этому аллелю, будет альбиносом независимо от состояния главного локуса.
2. Сцепление. В экспериментах по гибридизации, проводившихся Менделем, факторы, которые он изучал, распределялись независимо, однако теперь мы знаем, что гены расположены в хромосомах (см. разд. 2.2.1) и поэтому распределение каждого из них по гаметам зависит от их относительного распределения по хромосомам. Если два локуса находятся в одной и той же хромосоме и притом расположены близко один от другого, то они будут передаваться вместе.
Например, Томас Хант Морган (Thomas Hunt Morgan), положивший в 1909 г. начало генетическому изучению плодовой мушки (Drosophila), обнаружил у двух генов D. melanogaster отклонение от второго закона Менделя; один из этих генов определяет цвет глаз (рг — пурпуровый, рг+— красный), а дру-. гой — длину крыла (vg —зачаточное, vg+ — нормальное). Обратите внимание на то, что принцип обозначения генов здесь несколько иной: обозначение получает мутантный (обычно рецессивный) аллель, а доминантный аллель отличают от него индексом +. Такая система обозначений была принята потому, что для каждого локуса дрозофилы обнаружено большое число мутантных аллелей; она до сих пор используется генетиками, работающими с дрозофилой, наряду с другими обозначениями (см. (!)). Морган скрещивал гетерозиготных самок (ргрг+ vgvg+) с гомозиготными самцами (prpr vgvg). На основании классической менделевской генетики следовало ожидать, что в fi все четыре возможные сочетания генов будут представлены в равных количествах: prpr vgvg, prpr vgvg+, prpr+ vgvg, prpn v§v§+ (рис. 2.3, А). Однако на самом деле частота родительских форм (prpr+ vgvg+ и prpr vgvg) в потомстве была в 10 раз выше частоты двух других форм. Морган объяснил это, допустив, что два родительских аллеля сцеплены один с другим и поэтому передаются в одной родительской хромосоме, как это показано на рис. 2.3, Б. Но если эти два гена сцеплены, то как вообще могли возникнуть генные комбинации prpr vgvg+ и prpr+ vgvg? Очевидно, генетический материал каким-то образом переносится с одной хромосомы на другую (рис. 2.3, В), и существование такого явления, названного кроссинговером, было впоследствии подтверждено цитологическими наблюдениями. Ясно, что чем ближе друг к другу расположены два локуса в хромосоме, тем выше вероятность их совместной передачи и тем менее вероятен их кроссинговер. Таким образом, по частоте сцепления между локусами можно судить об их относительном местоположении в хромосомах, что служит основой для составления карт хромосом.

Рис. 2.3. Интерпретация механизма наследования цвета глаз и формы крыла у дрозофилы (объяснения см. в тексте).
Первым признаком, хромосомную локализацию которого Удалось установить (в результате ряда ключевых экспериментов, проведенных главным образом в США между 1900 и 1910 гг.), был пол. У млекопитающих и у дрозофилы одна хромосома (назовем ее хромосомой X) имеется у особей женского пола в двух экземплярах, а у особей мужского пола — только в одном. Вместо второй Х-хромосомы у самцов имеется отличная от нее Y-хромосома. Таким образом, самки несут хромосомы XX и образующиеся у них яйцеклетки содержат одну Х-хросому, а самцы несут хромосомы XY и образующиеся у них сперматозоиды содержат либо Х, либо Y-хромосому. Если допустить, что находящиеся в этих хромосомах ген или гены контролируют пол, то становится возможным объяснить наблюдаемое соотношение полов 1 : 1 при помощи следующей простой схемы:
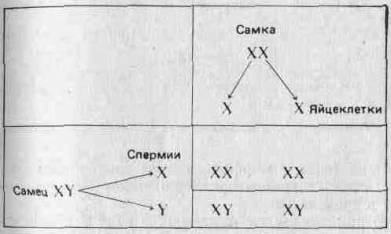
(Отметим, что у птиц и у некоторых насекомых наблюдается обратная картина: гетерогаметны самки, а самцы гомогаметны.) Соотношение полов рассматривается более подробно в разд. 3.7. Половые хромосомы X и Y морфологически отличаются от остальных хромосом (аутосом) и друг от друга. Поэтому можно ожидать, что они содержат уникальные аллели, которые не могут переходить из одной хромосомы в другую в результате кроссинговера. Это может привести к отклонениям от предсказаний Менделя; явление это известно под названием сцепления с полом. Например, аллель, определяющий красный цвет глаз У Дрозофилы, о котором говорилось выше, как и аллель белого Цвета глаз, локализован в Х-хромосомах. В Y-хромосомах этих аллелей нет. Красный цвет глаз доминирует над белым. Когда Морган скрещивал красноглазых самок с белоглазыми самцами, все потомство fj оказалось красноглазым. При скрещивании красноглазых самцов и самок из fi он получил красноглазых и белоглазых мух в соотношении 3:1, но все белоглазые мухи были мужского пола. Отношение красноглазых самок к красноглазым самцам составляло 2:1. Если, заменив обозначения, принятые на стр. 27, обозначить аллель красных глаз через Xw, а аллель белых глаз — через Xw, то нетрудно дать объяснение! наблюдаемым результатам:
Рациональное объяснение этого дает рис. 2.5. Вообще чем больше генов участвует в определении данного признака, тем более непрерывна изменчивость по этому признаку.
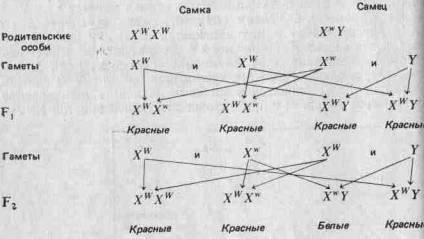
Такая ассоциация генов с морфологически различными хромосомами служит очень хорошим подтверждением того, что гены организованы в хромосомы.
3. Непрерывно изменяющиеся признаки. Мендель сконцентрировал внимание на признаках, резко отличающихся один от другого, однако большинству признаков свойственна непрерывная изменчивость. Так, например, организмы по общим размерам обычно не распадаются на дискретные классы, а образуют непрерывный ряд от минимальных до максимальных для данной группы размеров. Кроме того, чаще всего особи распределяются по данному признаку таким образом, что большая их часть концентрируется вблизи среднего значения признака, а по обе стороны от него частота постепенно сходит на нет (см., например, рис. 2.4). В течение некоторого времени полагали, что эта изменчивость выходит за рамки менделевской генетики; У. Ф. Уэлдон (W. F. Weldon) примерно в 1900 г. даже подверг критике данные самого Менделя, поскольку при более тщательном анализе оказалось, что большая часть так называемых дискретных признаков горошка подвержена значительной изменчивости (например, степень морщинистости семян и интенсивность их зеленой окраски). Однако в 1909 г. X. Нильсен-Эле (Н. Nilssen-Ehle) показал, что менделевские факторы могут контролировать количественные признаки, если один такой признак детерминируется большим числом генов (полигены), каждый из которых обладает небольшим, аддитивным действием (см. стр. 26).
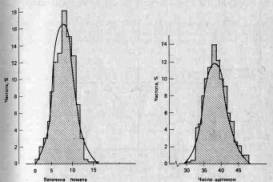
Рис. 2.4. Частотное распределение величины помета у мышей и числа щетинок на вентральной поверхности брюшка у дрозофилы. В обоих случаях кривая распределения приближается к нормальной (гауссовой) кривой. (Futuyama D. J., Evolutionary Biology, Sinauer Associates, Massachusetts, 1979.)
Помимо генетической основы изменчивости признаков существует еще и влияние окружающих условий. Например, на общие размеры животного или растения решающее влияние оказывают те условия, в которых оно развивается. Возможно, все признаки организма представляют собой продукт воздействия генетических и средовых факторов, так что нормальные распределения, подобные изображенным на рис. 2.4, — результат сочетания этих факторов. Передается по наследству только та компонента этой изменчивости, которая обусловлена генами; ее выражают как Долю общей изменчивости и называют наследуемостью (часто обозначается как h2). Измерение наследуемости имеет важное значение для селекционеров и животноводов; ее можно определить по соотношению между признаками родительских особей и потомков, выращенных в тех же условиях.
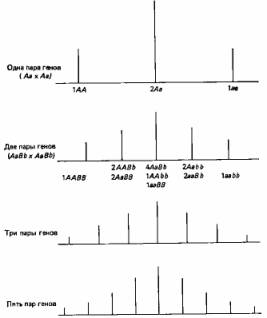
Рис. 2.5. Схема, иллюстрирующая возникновение фенотипической изменчивости при корпускулярной наследственности.
Допустим, что «сила» данного признака зависит от соотношения доминантных и рецессивных аллелей. В таком случае, чем больше число расщепляющихся локусов, определяющих этот признак, тем больше число возможных сочетаний доминантных и рецессивных признаков, а тем самым и число возможных фенотипических классов. (Strickberger M. W., Genetics, Macmillan, New York, 1968.)
Вильгельм Людвиг Иогансен (W. L. Johansen), специалист по генетике растений, был одним из первых ученых, обратив- J ших внимание (примерно в 1900 г.) на то, что каждый организм— продукт среды, в которой он обитает, и генов, которые он содержит. На этой основе Иогансен различал фенотип, то есть, организм, который мы видим, и генотип — совокупность содержащихся в нем генов, и это проведенное им разграничение сохранило свое важное значение по сей день.
4. Мейотшеский драйв. Аллели, кодирующие один и тот же признак, находятся в отдельных хромосомах (гомологичных хромосомах), так что каждая клетка данной родительской особи содержит обычно некоторое число пар гомологичных хромосом (диплоидное состояние). При формировании гамет гомологичные-хромосомы должны быть отделены одна от другой, с тем чтобы каждая гамета в конечном итоге получила по одной хромосоме от каждой пары (гаплоидное состояние). Этот процесс разделения известен под названием мейоза и лежит в основе первого закона Менделя — закона расщепления. Мейоз обеспечивает формирование гетерозиготной особью одинакового числа гамет, содержащих разные аллели. Однако некоторые процессы, детерминируемые генами, способны нарушить эти результаты. Такие процессы могут быть обусловлены событиями, как предшествующими мейозу, так и следующими за ним; и те и другие приводят к так называемому мейотическому драйву. Обзор по этой проблеме см. (16).
Один интересный пример связан с так называемыми генами SD (segregation distorter), обнаруженными у дрозофилы (5). В этом случае гомологичные хромосомы конъюгируют, но гены SD оказывают на своих партнеров такое воздействие, что те в дальнейшем образуют нежизнеспособные сперматозоиды. Существование подобных SD-генов проявляется более четко в тех случаях, когда они редки, потому что при этом у них больше шансов оказаться в гетерозиготном состоянии. По мере того как они становятся более обычными, конъюгация между ними происходит чаще и они разрушаются. Поэтому следует ожидать, что гены SD сбалансированы в популяциях на довольно низких уровнях.
2.2. Хромосомные и химические основы наследственности.
2.2.1. Цитологические основы наследственности.
За последние три десятилетия XIX в. в изучении клеточной основы жизни были достигнуты выдающиеся успехи. Ряд исследователей, главным образом немецких, открыли, что гаметы представляют собой клетки и что клетка — это не просто комочек однородного вещества, а некая структура, состоящая из ядра и цитоплазмы. Благодаря усовершенствованию микроскопов и разработке специальных методов окрашивания стало возможным более детально рассмотреть ядро. Оказалось, что оно также неоднородно: в нем были обнаружены нитевидные структуры, состоящие из вещества, названного хроматином; соответственно структуры эти получили название хромосом. Мнение о том, что хромосомы — подходящие кандидаты на роль материальных носителей наследственности, впервые высказал Август Вейсман (August Weismann, 1834—1914). Он представлял себе наследственность как передачу материала — того, что он называл зародышевой плазмой, — от родителей потомкам и предполагал, что зародышевая плазма, сохраняя непрерывность и обособленность, проходит через весь онтогенез данного индивидуума и «выплескивается» в следующее поколение, покидая сому — свое прежнее тело. Эти взгляды Вейсмана были основаны на его наблюдениях, которые показывали, что у многих животных образование половых клеток и их обособление от соматических тканей происходит на очень ранней стадии эмбрионального развития. Отсюда проводившееся им разграничение между зародышевой плазмой и сомой. В своей «Эволюционной теории», вышедшей в 1903 г., Вейсман еще более сузил границы зародышевой плазмы и свел ее к ядру половых клеток, а затем — к хромосомам и хроматину (впервые эти термины были предложены В. Вальдейером в 1888 г.). Он это сделал потому, что, судя по гистологическим данным, хромосомы вели себя именно так, как им следовало себя вести, если бы они представляли собой вещество наследственности: они удваивались и разделялись на две равные группы при делении соматических клеток; число их уменьшалось вдвое при образовании гамет, предшествующем смешиванию мужского и женского вкладов во время оплодотворения. В настоящее время мы называем эти два процесса соответственно митозом и мейозом.
Итак, на основе всех полученных данных менделевские факторы получили название генов (термин, введенный В. Л. Иоган-сеном в 1888 г.), которые, как предположили исследователи, находятся в хромосомах. Эта теория, конечно, коренным образом отличалась от теории слитной наследственности, которой придерживался Дарвин. Зародышевая линия была отделена от сомы, а тем самым и от приобретенных признаков. Гены дискретны и не сливаются. Кроме того, хромосомная теория позволяла лучше осмыслить данные некоторых экспериментов по скрещиванию, например данные Моргана по сцеплению у дрозофилы (см. разд. 2.1.2).
2.2.2. Химические основы наследственности.
Локализация генов в хромосомах была установлена, однако не все биологи были так уверены, как Вейсман, в их чисто химической природе. Сам Т. Г. Морган в своей книге «Механизм менделевской наследственности» (Т. Н. Morgan, A. H. Sturte-vant, H. J. Muller, С. В. Bridges, Mechanism of Mendelian Heredity, H. Holt and Co., 1915) писал: «Предположение, что частицы хроматина, неотличимые одна от другой и почти гомогенные при исследовании любыми известными методами, могут в силу своей материальной природы наделять всеми свойствами жизни, превосходит воображение даже самого убежденного материалиста».
Создание разнообразия, или, иными словами, генетической информации, из единообразия —вот в чем заключался главный вопрос, а ответ на него состоял в том, что хроматин оказался не таким гомогенным, каким он представлялся. Данные хими-деского анализа показали, что хроматин — это нуклеиновая кислота, состоящая из повторяющихся субъединиц (нуклеоти-дов), соединенных друг с другом связями, в образовании которых участвуют остатки либо рибозы (рибонуклеиновые кислоты, РНК), либо дезоксирибозы (дезоксирибонуклеиновые кислоты, ДНК).
Крупнейший шаг вперед в этой области был сделан в середине пятидесятых годов нашего века, когда Дж. Д. Уотсон (James D. Watson) и Фрэнсис Крик (Francis Crick) расшифровали строение ДНК на основе химических данных в сочетании с рентгеновскими «снимками» ее молекулы. Каждая нуклеиновая кислота состоит из двух цепей, соединенных химическими связями, так что структура в целом имеет вид веревочной лестницы. Перекладинами этой лестницы служат связи между нуклеотидами, причем нуклеотиды расположены таким образом, что против определенной большой единицы может находиться только одна определенная малая единица. Поэтому говорят, что две стороны лестницы комплементарны. Эта лестница свернута наподобие гибкого электрического провода, образуя ныне знаменитую двойную спираль (см. рис. 2.6).
Вслед за созданием модели структуры ДНК был высказан ряд соображений относительно ее функций. Во-первых, последовательность оснований в молекуле ДНК может быть различной и в этой последовательности может быть закодирована генетическая информация. Хроматин оказался негомогенным. Во-вторых, комплементарность означает, что если отделить две стороны «веревочной лестницы» одну от другой, то каждая из них могла бы служить матрицей для построения другой. Следовательно, информация может точно и безошибочно реплицироваться и передаваться от одной клетки другой и от родителей потомкам (рис. 2,6,Г).

Рис. 2.7. Транскрибирование ДНК в мРНК и трансляция в белки. Трансляция происходит на субклеточных органеллах — рибосомах; молекулы транспортной РНК (тРНК) выбирают из клеточного фонда аминокислот нужную аминокислоту и переносят ее на соответствующее место на матрице. Образующийся таким образом белок может затем выполнять в клетке структурные или ферментативные функции. (Calow P., Biological Machines, Edward Arnold Publ., London, 1976.)
Наконец, спиральная структура означает, что эта жизненно важная генетическая информация защищена от превратностей внешнего мира. Время от времени в процессе репликации возникают случайные нарушения, ведущие к тому, что в классической генетике называли мутациями. Подобные события должны быть редкими и могут вызываться внешними факторами (например, облучением), но они не направлены на удовлетворение каких-либо определенных «потребностей».
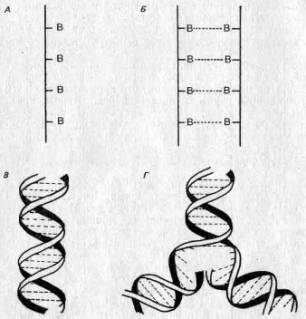
Рис. 2.6. Строение ДНК. А. Нуклеотиды (В), соединенные в цепь. Б. Комплементарные цепи, образующие «веревочную лестницу» при помощи водородных связей (прерывистые линии). В, «Веревочная лестница», свернутая в двойную спираль. Г. Репликация: цепи раскручиваются, и каждая из них служит матрицей для образования комплементарной цепи.
Все эти положения, разработанные Уотсоном и Криком, оказались в основном верными и вызвали бурное развитие исследований по молекулярной генетике. Было известно, что ядро и нуклеиновые кислоты играют ключевую роль в синтезе белка, а поскольку белки принимают важнейшее и непосредственное участие в построении клетки (в виде строительных блоков) и в регуляции ее функций (в виде ферментов), то нетрудно видеть, каким образом нуклеиновые кислоты могли бы определять признаки организма. Нуклеиновые кислоты состоят из повторяющихся субъединиц — нуклеотидов, а белки состоят из аминокислот; поэтому можно было допустить, что последовательность нуклеотидов кодирует последовательность аминокислот. Поскольку, однако, аминокислот различных типов в белках больше, чем нуклеотидов в нуклеиновых кислотах (соотношение их составляет примерно 5:1), одна аминокислота не может кодироваться только одним нуклеотидом. Теоретически было нетрудно показать, что лишь группы из трех нуклеотидов (триплеты) могут обеспечить достаточное разнообразие (43, или 64 альтернативные комбинации), а вскоре существование три-плетного кода было установлено экспериментально. У эукариот перенос генетической информации при синтезе белка происходит в два этапа: от дезоксирибонуклеиновых кислот (ДНК) к рибонуклеиновым кислотам (РНК)—процесс, известный под названием транскрипции, и от РНК к белкам — трансляция. Упрощенная схема переноса генетической информации представлена на рис. 2.7.
2.2.3. Связь классической и молекулярной генетики.
Какая часть ДНК или какое ее количество эквивалентно одному менделевскому гену? Задать такой вопрос естественно, но ответить на него невозможно. Во-первых, следует отметить, что в нуклеиновых кислотах закодированы не форма носа, цвет волос или другие подобные признаки, а молекулы, необходимые для создания и поддержания этих признаков! Во-вторых, с точки зрения молекулярной конституции организма подразделение его на признаки, принятое в генетике, весьма произвольно. Например, какая-либо одна молекула, образованная одной частью ДНК, может содержаться в носе, волосах и глазе. Это с неизбежностью привело к тому, что концепцию гена оказалось необходимым пересмотреть, определив его в терминах дискретных частей генома, то есть тех его частей, которые продуцируют конкретные белки. Однако не все гены кодируют синтез белков; некоторые из них регулируют активность других генов, а поэтому первые получили название структурных генов, а вторые — генов-регуляторов. Примерно 30 — 70% ДНК млекопитающих представлено повторяющимися участками, и если они вообще несут какую-либо функцию, то, вероятно, эта функция регуляторная (см., однако, разд. 3.8). Наконец, начинают появляться данные о том, что участки ДНК, кодирующие определенные белки, не обязательно физически дискретны и статичны. Гены и фрагменты генов могут перемещаться в молекуле ДНК, и такие подвижные гены были обнаружены как у микроорганизмов, так и у эукариот. Менделевская концепция физически дискретных (корпускулярных) генов становится менее определенной.
Связь между ДНК, менделевскими факторами и признаками организма не столь проста, как представлялось прежде, но под наблюдения классических менделистов можно тем не менее подвести химическую основу. Установленные ранее явления доми-нантности и рецессивности рассматриваются теперь в таких аспектах, что можно без труда показать связь этих явлений с определенными химическими реакциями (разд. 2.1.2). Эпистаз (разд. 2.1.2) можно интерпретировать в понятиях взаимодействующих генов и генных продуктов. Нетрудно, например, представить себе, как может происходить химическое воздействие на экспрессию на уровне как самого гена, так и любого из ряда последовательных генных продуктов химическими факторами, содержащимися во внутренней и во внешней среде. Наконец, мутацию можно интерпретировать как ошибку в репликации ДНК. Такая ошибка может ограничиваться замещением одного нукле-отида другим (точковая мутация) или же выражаться в структурной перестройке целых хромосом (хромосомные мутации, в том числе нехватки, удвоения, транслокации и инверсии).
2.2.4. Снова ламаркизм?
Даже до того, как была полностью расшифрована структура нуклеиновых кислот, стало ясно, что информация должна передаваться в направлении ДНК—Мэелки, а не наоборот. Это положение, известное как «центральная догма молекулярной биологии», соответствует представлениям Вейсмана о том, что генетическая информация передается в одном направлении — от зародышевой плазмы к соме. Однако, несмотря на то что центральная догма' и концепция Вейсмана совместимы, между ними все же есть различие. Первая запрещает поток информации от цитоплазмы клетки к ее ядру, а вторая — от клеток одного типа (соматических) к клеткам другого типа (зародышевой линии). Обе отрицают ламаркизм. Но если центральная догма нерушима, вейсманизм уязвим, по крайней мере в принципе. Известны, например, вирусы, способные переносить нуклеиновые кислоты из одной бактериальной клетки, выращиваемой в культуре, в другую такую клетку, и это, в сущности, составляет один из основных методов генетической инженерии. Нет причин, в силу которых не могла бы происходить такая же передача информации от клетки к клетке в многоклеточных организмах, а раз так, то почему информация не может передаваться от соматических клеток половым? Исходя из этих соображений Э. И. Стеель в своей книге «Соматический отбор и приспособи-тельная эволюция» (Е. J. Steele, Somatic Selection and Adaptive Evolution, Croom Helm, 1980) недавно вновь обратился к ламаркизму.
Известно, что в соматических клетках иногда возникают случайные мутации. Полезные мутации, то есть мутации, благоприятные для несущей их клетки, могут распространиться, так как они дают возможность своим носителям делиться быстрее, чем другие клетки; это в особенности относится к клеткам, участвующим в борьбе с инфекцией. Чем больше они преуспевают, тем более многочисленными они могут оказаться. Таким образом, мутантный ген размножается, а при этом повышаются его шансы на то, что он будет захвачен вирусами и перенесен в другие клетки, возможно в том числе и в клетки зародышевой линии. По-видимому, у любого индивидуума существует период отбора соматических мутаций, предшествующий прохождению мутаций сквозь фильтр дарвиновского естественного отбора. Стеель считает, что этот процесс должен ускорять эволюцию и позволяет легче объяснить эволюцию таких сложных и координированных органов, как глаз.
Экспериментальных данных, которые бы подтверждали эти идеи, пока что немного, но они, во всяком случае, правдоподобны. Вместе с тем эта теория сталкивается с рядом трудностей.
Во-первых, не все клетки организма способны размножаться — это относится, в частности, к нервным и мышечным клеткам, так что соматический отбор, очевидно, возможен не во всех случаях. Во-вторых, распространение мутаций среди соматических клеток нельзя автоматически расценивать как доказательство их полезности. Именно так ведут себя онкогенные мутации, однако они летальны. Но если даже окажется, что идеи Стееля верны, то ничего таинственного в них нет, и нет никаких причин возрождать таинственное стремление к удовлетворению неких особых потребностей в качестве основы для объяснения разнообразия живых существ и их способности к адаптации. В сущности, можно сказать, что гипотеза Стееля все еще опирается главным образом на случайную изменчивость и естественный отбор, а сделанный им вклад сводится к тому, что отбор происходит не только между организмами, но и внутри них. Еще в 1881 г. Вильгельм Ру в своей книге «Борьба частей в организме» (Wilhelm Roux, Der Kampf der Theile im Organismus, Leipzig) также высказал предположение, что в организме, возможно, происходит борьба (а следовательно, отбор) между тканями и органами за ограниченные ресурсы, получаемые с пищей, и что она может оказаться важной силой в эволюции. По иронии судьбы, Вейсман также признавал возможность внутриорганизменного отбора, считая, что он может играть важную роль в морфогенетической пластичности, но затем отбросил его как несущественный элемент эволюции.
2.3. Неодарвинизм и популяционная генетика.
Разрешив отдельные проблемы, с которыми столкнулась теория Дарвина, менделизм вместе с тем породил и ряд новых. Во-первых, стали высказываться предположения, что такие генетические явления, как доминирование, возможно, играют более важную роль в определении признаков популяции, чем сам естественный отбор. Ведь проводимые в лаборатории скрещивания между особями, гомозиготными по доминантным и особями, го-мозиготными по рецессивным генам, приводят к численному превосходству доминант (см. разд. 2.1.1). Во-вторых, дискретная, корпускулярная природа наследственных факторов должна означать, что изменения этих факторов также дискретны и что каждое изменение, по всей вероятности, будет иметь значительный эффект. Изменчивость, следовательно, носит характер больших скачков, а не мелких непрерывных изменений, как это представлял себе Дарвин.
Де Фриз в своей книге «Мутационная теория» (De Vries, The Mutation Theory, Open Court Publ. Co., 1909) проводил различие между непрерывной индивидуальной изменчивостью и дискретными скачкообразными изменениями. Он применял термин- мутация только к последним (однако, как должно было стать очевидным из всего сказанного выше, этот термин применяется теперь в более общем смысле для обозначения любого спонтанного случайного изменения, большого или малого). Как считал де Фриз, именно эти мутации контролируют эволюционное изменение и играют более важную роль, чем сам естественный отбор.
Ранние менделисты, подобно де Фризу, преуменьшали эффективность естественного отбора и подчеркивали первостепенное значение менделевских соотношений и мутаций в формовке эволюции. На некоторых дарвинистов это произвело такое впечатление, что они начали высказывать сомнения в обоснованности менделизма, и между этими двумя группами начались ожесточенные дебаты. Другие эволюционисты, понимая важность менделизма и таящиеся в нем возможности, вознамерились примирить его с дарвиновской теорией. Поскольку возникшая в результате их усилий синтетическая теория эволюции выходила за рамки механизмов, предложенных Дарвином, ее часто называют неодарвинизмом. Немалую роль в решении этой проблемы сыграл переход от рассмотрения последствий скрещивания между двумя особями к последствиям скрещивания между особями в популяциях; менделевские соотношения превратились, таким образом, в частоты различных генотипов в дайной популяции, а генетические и эволюционные изменения выражались в изменениях частот соответствующих генов. В результате генетика превратилась в популяционную генетику, которая развивалась параллельно с неодарвинизмом. В дальнейших разделах настоящей главы дан обзор основных моментов этого развития л показано, как популяционная генетика пыталась ответить на ключевые вопросы, поставленные менделистами.
2.3.1. Уравнение Харди—Вайнберга
В 1908 г. Дж. Харди (G. H. Hardy) и В. Вайнберг (W. Wein-berg) независимо друг от друга занялись изучением следующего вопроса, поднятого менделистами: могут ли изменения генных частот происходить под влиянием одних только ограничений, налагаемых законами Менделя? Для того чтобы ответить на этот вопрос, они представили себе популяцию, в которой происходит случайное скрещивание (панмиксия) и численность которой достаточно велика, чтобы полученные результаты можно было считать статистически достоверными. Отметим, что в подобной ситуации (в отличие от экспериментов Менделя) нельзя точно знать, какие именно будут происходить скрещивания, однако можно строить предположения о вероятности тех или иных скрещиваний на основе частоты особей, несущих определенные гены. Харди и Вайнберг допустили, что в их воображаемой популяции имеются два аллеля, Л и а, с первоначальными частотами соответственно р и q, так чт p + q= (то есть это условие позволяет описать все возможные случаи, поскольку в данном локусе нет других аллелей). При этом могут образоваться следующие дишюидные генотипы: АА, Аа и аа. Харди и Вайнберг допускали свободное скрещивание, так что вероятность встречи двух А-гамет и образования особи АА равна р2. Это следует из основной теоремы теории вероятностей, согласно которой вероятность совместного появления двух последовательных событий (скажем, выпадения два раза подряд орла при бросании монеты) равно произведению вероятностей каждого из них, то есть
вероятность выпадения орла равна ~- •_-, или (g-)2. Рассуждая
подобным же образом, получим, что вероятность aa=q2, а вероятность Aa=pq. Согласно правилу Менделя, частота гетерозигот должна быть вдвое выше частоты каждой из гомозигот, так что ожидаемые частоты будут следующими: p2 + 2pq + qz. Частота гена А в новом поколении выражается формулой (p2+pq)/(p2 + + 2pq + q2), которую можно преобразовать к виду (p(p + q))f J(p + q)2, а поскольку p + q = l, она сводится к р. Однако общая частота генов в этом локусе равна, согласно определению, 1, так что частота " аллелей а составляет (1 — р), то есть q (опять-таки по определению). Итак, частоты генов остались в точности прежними и будут продолжать оставаться такими в каждом из последующих поколений. Следовательно, менделевские механизмы сами по себе не вызывают изменений в частотах генов и поэтому не могут быть важным фактором, направляющим эволюционное изменение. Очевидно, в этом должны участвовать другие факторы.
2.3.2. Холдейн, Фишер и значение отбора
Может ли естественный отбор распознавать мелкие различия в генетически детерминированных признаках, оказывающие лишь небольшое воздействие на выживание и размножение их носителей? Могли ли эти мелкие случайные различия сами по себе вызывать существенные изменения? Или же нам следует допустить, что основным материалом для эволюции служат крупные скачкообразные изменения? Эти вопросы поставили Дж. Б. С. Холдейн в своей книге «Генетическая теория эволюции» (J. В. S. Haldane, Genetlcal Theory of Evolution, Longmans, 1930) и Р. А. Фишер в «Факторах эволюции» (R. A. Fisher, The •Causes of Evolution, Oxford, 1932). Эти две книги сыграли чрезвычайно важную формирующую роль в дальнейшем развитии неодарвинизма. Еще одной важной фигурой в его развитии был Сьюэлл Райт (Sewall Wright). Однако, подход Райта к этим вопросам настолько отличался от позиций Холдейна и Фишера, что мы рассмотрим его отдельно в разд. 2.3.3.
И Фишер, и Холдейн выражали интенсивность отбора через изменения в частотах генов. Те аллели, частота которых относительно других аллелей выше, считаются более приспособленными, поэтому изменение относительных частот аллелей служит мерой «неодарвиновской» приспособленности. (Обратите внимание, что нельзя ограничиваться рассмотрением абсолютных изменений, так как при понижении абсолютной плотности данного аллеля в популяции он все же может оставлять больше своих представителей, чем другие аллели, если плотность этих других аллелей уменьшается быстрее. В таком случае он все еще более приспособлен, поскольку в преобладающих в данной среде условиях он по сравнению с другими аллелями оказался более преуспевающим.)
Проще всего начать анализ с динамики генов в гаплоидных популяциях (то есть в популяциях, в которых каждая особь содержит по одному аллелю на локус); в табл. 2.2 представлены основные расчеты для двух аллелей (Л и а) в такой популяции. Холдейн измерял приспособленность, сравнивая чистые скорости прироста R, то есть делил значение R для каждого аллеля на максимальное значение R, так что приспособленность наиболее преуспевающего аллеля становится равной 1, а приспособленность других аллелей — какой-то доле 1. Этот индекс обычно обозначают буквой w и называют селективной или адаптивной ценностью. В некоторых случаях эту величину можно записать в виде 1 —w ( = s), что выражает степень, в которой данный ген подвергается отрицательному отбору; s называют коэффициентом отбора.
Таблица 2.2. Вычисление приспособленности в гаплоидной популяции
| Аллель | Численность в одном поколении | Численность в одновременном следующем поколении | Чистый прирост за период между нулевым временем и временем t | Показатель приспособленности ' Холдейна (относительно Л) |
| А | N0 | Nt | Nt --- = RA N0 | 1 |
| а | N0 | nt | nt ---- = Ra n0 | Ra/RA |
Это крайне упрощенный пример. Время генерации у аллелей А и а может быть различным, и в таком случае придется вносить поправку путем деления всех величин на время генерации. Если поколения перекрываются (то есть особи, составляющие популяцию в данный момент времени, имеют разный возраст) и размножение у разных поколений происходит несинхронно, то положение еще более осложняется, потому что в этом случае нет стандартных интервалов, которые можно было бы использовать для сравнения изменений частоты аллелей А и а. Поэтому Фишер предпочел использовать в качестве меры приспособленности относительные скорости размножения генов в данной популяции, так как в экстремальном случае непрерывного размножения (какие-то особи из тех или иных поколений приносят потомков все время) эту скорость можно вычислить из изменений абсолютной численности (например, N для аллеля А и п для аллеля а) на протяжении какого-либо промежутка времени. Подобный мультипликативный рост описывается уравнениями dN/dt—rAN и dn/dt = ran, которые после интегрирования приобретают вид Nt = N0erAt и п( = поега( (е — основание натуральных логарифмов, r — константа, t — время, А к а — аллели, к которым относится данный параметр). Коэффициент г, который Фишер назвал «мальтузианским параметром» (поскольку Мальтус был одним из первых, создавших модель мультипликативного роста), представляет собой меру скорости распространения каждого гена и поэтому служит показателем приспособленности. Обратите внимание, что если мы примем t за среднее время генерации, как в случае прерывистого роста, то тогда
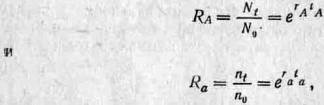
и если, из соображений удобства, мы примем tA = ta=, то RalRA = era~-rAn поэтому In wa=ra—га- Следовательно, эти два показателя приспособленности связаны друг с другом. Популицонные генетики обычно предпочитают пользоваться холдейновской мерой приспособленности, тогда как эволюционные экологи часто используют меру, предложенную Фишером. В этом разделе мы в дальнейшем будем пользоваться параметрами w и s, а к коэффициенту r вернемся в разд. 3.4. (Более подробное рассмотрение коэффициента r и его ограничений в качестве меры приспособленности можно найти у Чарлзуорта (3).)
Уравнения для диплоидного случая гораздо сложнее, чем для гаплоидного, потому что степень распространения или сокращения данного аллеля в популяции зависит от того, с какими аллелями он становится связанным в гетерозиготах. Так, например, вредный рецессивный ген будет защищен от элиминации отбором, если он в гетерозиготном состоянии связан с благоприятными доминантными аллелями. И напротив, отбор, направленный против вредного доминантного аллеля, ведет к элиминации благоприятных рецессивных аллелей, если они связаны с ними в гетерозиготах.
Приспособленность генотипов можно вычислить в основном таким же способом, как и приспособленность составляющих их аллелей. Это довольно искусственный способ, потому что гомо-зиготы одного поколения могут вносить вклад в гетерозиготы другого и наоборот (то есть поставляются последующим поколениям гены, а не генотипы). Тем не менее из этих частот генотипов можно вывести изменение частоты аллелей при условии, что мы располагаем дополнительной информацией: а) о начальных частотах аллелей и б) о степени доминирования. Мы делаем это, используя адаптивные ценности и коэффициенты отбора, а не мальтузианские параметры, однако аналогичным образом можно использовать и последние (9).
Простейший возможный случай относится к одному локусу с двумя аллелями, например Л и а, с частотами соответственно р и q. Большая часть рассуждений в популяционной генетике основана на системе именно такого типа. Допустим, что в результате отбора равновесие Харди—Вайнберга в некоей воображаемой популяции сдвигается следующим образом:
Изменение, происшедшее с каждым генотипом (то есть эквивалент R в табл. 2.2), составит поэтому АА = 0,32/0,16 = 2; Аа — = 0,52/0,48=1,08; аа = 0,16/0,36=0,44; на основе этого можно получить значение приспособленности для каждого генотипа, разделив каждое из полученных чисел на 2 (то есть на самое большое значение).
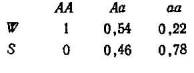
Отсюда (W и S набраны прописными, так как они обозначают генотипы, а не свойства генов; этого мы будем придерживаться по всей книге). Возвращаясь к начальным данным, можно получить новые частоты генотипов, установившиеся после отбора, из произведения исходных частот и значений W, приведя каждое из них к долям единицы путем деления на сумму этих отдельных членов. Эту сумму иногда обозначают через W, и она равна средней, по всем W.

В табл. 2.3 эти результаты представлены в обобщенном виде.| Из первоначальных частот генов и значений W мы можем теперь! вычислить конечные частоты аллелей (то есть q' и р'). Отсюда час-| тота аллеля а после отбора будет равна
Изменение частоты (А2) за одно поколение равно поэтому q'—q. Приведенные выше уравнения обычно записывают не через W, a через 5, но это лишь обычная замена переменных. Аналогичным образом р' — это просто 1 — q'.
Таблица 2.3. Обобщение результатов вычисления изменений в частотах генотипов, рассмотренных в тексте
| АА | Аа | аа | Всего | |
| Частота до отбора | Р2 | 2рд | 2pq | 1 |
| Приспособленность | Waa | Waa | Waa | |
| Пропорциональный вклад | Waap2 | Waa2pq | Waaq2 | W |
| Частота после отбора в одном поколении | Waap2 W | Waa2pq W | Waaq2 | 1 |
Скорость и характер изменения генных частот от поколения к поколению зависят: 1) от степени доминантности или рецессивности данного аллеля (которая изменяет соотношение значений: W, то есть при доминировании waa= WAa =/= Waa, а в отсутствие доминирования или при сверхдоминировании Waa =/= waa =/= Wqq).
2) от того, какому аллелю благоприятствует отбор (например как уже говорилось, вредный рецессивный аллель может быть скрыт в гетерозиготе, что замедлит его элиминацию; или же благоприятный рецессивный аллель в гетерозиготном состояние будет элиминироваться отбором, что задержит его распространение в популяции); 3) от интенсивности отбора. Использование этих величин приводит к тому, что представленные выше уравнения для изменения частот аллелей приобретают другой вид; вывод всех этих уравнений можно найти в большинстве учебников по популяционной генетике. Однако решения этих уравнений нельзя найти аналитически (то есть исходя из алгебраически уравнений); их можно получить только подстановкой конкретных вещественных чисел вместо q и W (то есть моделированием). Результаты, полученные на одной такой модели, действительно построенной Холдейном, приведены в табл. 2.4. Они относятся к случаю, в котором доминантный аллель обладает преимуществом при относительно низком давлении отбора. Это показывает, что изменение частоты генов зависит от их исходной частоты. При очень низких частотах отбор действует медленнее, чем при промежуточных, а по мере приближения данного аллеля к закреплению (то есть вытеснению другого аллеля) скорость изменения его частоты заметно снижается. Это происходит потому, что менее благоприятный аллель сохраняется в гетерозиготах и его трудно вытеснить полностью. Рецессивному аллелю, обладающему преимуществом, закрепиться при низких частотах еще труднее, поскольку, находясь в гетерозиготном состоянии, он элиминируется из популяции отбором, направленным против менее благоприятного доминантного аллеля, однако закрепление такого рецессивного аллеля происходит быстрее. Ряд моделей, в которых 5> или равно 0,001 и в которых рассматривались разные соотношения между Wгомоз. и Wгетероз., показывает, что сдвиги в генных частотах благоприятных аллелей от 0,1% до более чем 50% особей данной популяции происходят за менее чем 104 поколений. К этому числу мы еще вернемся.
Таблица 2.4. Число поколений, необходимых для того, чтобы произошло да! ное изменение в частоте доминантного аллеля, если доминант оставляют 1000 потомков на 999 потомков рецессивов, т. е Waa = 0,999 или Saa = 0,01
| Р Число поколений | 0,001 до 1% 6920 | 1 до 50% 4819 | 50 до 99% 11664 | 99 до 99,999' 309780 |
Если под действием отбора частота благоприятного аллел возрастает, то отбор должен также повышать среднюю приспособленность популяции в целом (то есть W). Кроме того, поскольку изменение частоты происходит медленнее всего, когда частота одного аллеля очень низка или высока, изменение это, вероятно, будет наибольшим при промежуточных частотах, то есть когда значения частот аллелей наиболее близки друг к другу. Это интуитивная формулировка принципа, который был строго доказан Фишером и считался основной чертой эволюции путем естественного отбора. Его часто называют основной теоремой естественного отбора Фишера, которая формулируется следующим образом: «скорость повышения приспособленности любого организма в любое время равна его генетической дисперсии по приспособленности в это время». Формулировка Фишера неудачна, так как под «организмом» он несомненно имел в виду популяцию. Генетическая дисперсия по W тем больше, чем ближе частоты аллелей и, следовательно, разброс значений W.
Фишер называл свою теорему основной, потому что она строго определяла зависимость между двумя краеугольными камнями дарвиновской теории — изменчивостью внутри популяций и скоростями эволюционного изменения. Однако эта зависимость оказалась не столь несомненной, как полагал Фишер. Так, например, если приспособленность сама зависит от генетической дисперсии (что представляется правдоподобным в некоторых ситуациях; см. разд. 3.5), то скорость изменения приспособленности не должна быть связана с этой дисперсией простой зависимостью. Кроме того, Фишер вывел свою зависимость для случая одного локуса с двумя аллелями; если же рассматривать примеры с участием более чем одного локуса, то зависимость между этими двумя переменными может оказаться значительно сложнее (см. разд. 2.3.3). Есть также и другие потенциальные неясности и проблемы (11). Таким образом, вытекающее из основной теоремы Фишера следствие, что действие естественного отбора всегда направлено на максимизацию приспособленности, нельзя принимать безоговорочно, и из этого в свою очередь вытекает ряд следствий, которые необходимо учитывать при изучении адаптации (см. гл. 3). С основной теоремой Фишера связана еще одна концепция — концепция генетического груза (различие между максимальной потенциальной приспособленностью популяции и W). Это различие создается любым процессом, порождающим генетическую изменчивость, в том числе мутационным процессом (мутационный груз) и процессом генетической перестройки, обусловленной расщеплением (сегрегационный груз). Как и основная теорема Фишера, а возможно, именно из-за нее, концепция генетического груза небесспорна и вызывает разногласия, а поэтому в дальнейшем мы о ней говорить не будем.
Вернемся теперь к вопросам, поставленным в начале этого раздела: насколько отбор важнее, чем повторные мутации? Обратите внимание на то, что для повышения частоты данного аллеля в результате мутирования при отсутствии отбора необходимо, чтобы данная мутация время от времени возникала вновь, то есть повторялась. Рассмотрим мутирование в одном направлении от Л к а, происходящее с частотой и. Вообще обычно происходит также обратное мутирование от а к А, уравновешивающее эффект первого процесса. Однако мы пренебрежем обратными мутациями и используем наиболее благоприятную систему для изменения частоты аллеля путем мутаций. Согласно оценке Холдейна, сделанной на основе наблюдений по дрозофиле и примуле, частота мутаций равна примерно 10~6 на один ген; эта оценка была подтверждена в дальнейшем исследованиями, проводившимися на других организмах. Холдейн вычислил потенциальное воздействие мутаций следующим образом: частота аллеля А в первом поколении потомков (pi) равна (1—и)р<> (где ро — исходная частота Л), а во втором поколении потомков (р2) она равна (1 — и)р! или (1 — и)2р0 и так далее. Записав эти формулы в общем виде, pt=( — и)*р0, и разделив правую и левую части на ро, а затем прологарифмировав, получим
Теперь мы можем использовать это уравнение для выяснения того, сколько потребуется поколений при частоте мутаций 1:106, чтобы сдвинуть частоту аллеля а (то есть д) с 0,1 до 50%. Если р=—д, то, следовательно, частота аллеля А должна при этом измениться от 99,9 до 50%; поэтому
Вспомним, что соответствующее значение для действия слабого отбора составляло менее 104 поколений. Таким образом, даже при самых благоприятных допущениях для мутационного процесса и неблагоприятных для естественного отбора последний оказывается гораздо более мощным (почти на два порядка) фактором изменения, чем первый.
Конечно, при этом все еще остается открытым вопрос относительно важности крупных мутаций по сравнению с мелкими. Фишер, как и Дарвин (см. разд. 1.5), утверждал, что вероятность ценности любой мутации обратно пропорциональна ее величине, поскольку вряд ли можно ожидать, что значительное резкое изменение окажется совместимым с другими признаками, с которыми оно должно образовать единое целое. К этому вопросу мы вернемся в гл. 5.
2.3.3. Сьюэлл Райт, адаптивные ландшафты и случай.
Приведенные выше анализы были основаны на рассмотрении пар аллелей, находящихся в одном и том же локусе. Однако большая часть фенотипических признаков контролируется совокупностями генов с участием многих аллелей и локусов. Эти гены могут быть в разной степени сцеплены друг с другом; кроме того, между ними могут происходить различные взаимодействия (см. разд. 2.1.2). Следовательно, приспособленность определенных аллелей в одном локусе зависит от аллелей, лежащих в дру* гих локусах, и, в сущности, на нее оказывает влияние вся генетическая среда, в которой они находятся. Быть может, группы сцепления, целые хромосомы или даже геномы представляют собой более значимые единицы отбора, чем отдельные гены (см. разд. 3.8). К сожалению, генетика таких полигенных систем чрезвычайно сложна, а поэтому мы не можем охватить ее в такой мере, как случай двух аллелей, лежащих в одном локусе. Мы исходим из допущения, что основные принципы остаются теми же, даже если существуют различия в деталях. Однако здесь возможны серьезные исключения.
Другой основатель неодарвинизма, Сьюэлл Райт (Sewall Wright), разработал концептуальную систему, позволяющую рассмотреть взаимодействие между генами и их влияние на приспособленность (см. Wright, 1931, Genetics, 16, 99—159). Она получила название адаптивного ландшафта или пространства приспособленности. В этой системе каждая из осей я-мерного пространства определяет частоту каждого гена в данной попу-ляции,_причем последняя ось определяет среднюю приспособленность W, так что каждое сочетание генов определяет некую частную приспособленность. Так, в простейшей из возможных трехмерной модели две оси соответствуют частоте двух аллелей в каждом из двух локусов, а пространство между этими осями — всем возможным сочетаниям этих генов. Третья ось соответствует значениям W для каждого сочетания. Итак, мы имеем трехмерный ландшафт (более сложные ситуации порождают п-мер-ный ландшафт, с которым столь же трудно иметь дело, как и просто представить его себе). Пример адаптивного ландшафта изображен на рис. 2.8. В этой модели наиболее приспособленная популяция находится на вершине «холмов», а действие отбора направлено на то, чтобы заставить популяции подниматься на вершины холмов и закрепляться на них. Не все адаптивные пики будут или должны быть заняты, поскольку число потенциально возможных сочетаний аллелей у одного вида больше числа самих видов. В связи с этим многие пики ни разу не будут «опробованы» и некоторые из них будут выше чем другие. На какой пик поднимутся популяции, зависит от близости популяций к этим пикам и, следовательно, от исходных генетических условий до начала отбора. В этом теория Райта существенно отличается от модели «два аллеля — один локус», которая предсказывает, что W всегда будет максимальным (то есть популяции должны подниматься только на самые высокие пики), а из того обстоятельства, что это не есть непременное условие, вытекает ряд следствий, имеющих значение для вопросов, относящихся к адаптациям организмов, которые мы рассмотрим в гл. 3.

Рис. 2.8. Адаптивный ландшафт при различных частотах двух хромосомных инверсий у одного из представителей прямокрылых. BL — инверсия Blundell; TD —инверсия Tidbinbilla. (Shorrocks В., The Genesis of Diversity, Hodder a. Stoughton, London, 1978, по данным Lewontin R. C., White M. J. D., Evolution, 1960, 13, 561—564.)
На рис. 2.9 адаптивные ландшафты использованы для того, чтобы представить в качественном аспекте некоторые возможные эволюционные ситуации. (Следует помнить, что мы рассматриваем эти ландшафты сверху, то есть видим их контуры.) На рис. 2.9, А показано, чего следует ожидать в случае ослабления действия отбора или усиления мутационного процесса: область, занимаемая некой популяцией, распространяется по склонам ее адаптивного пика, и если это происходит достаточно интенсивно, то вид, к которому она принадлежит, будет вынужден занять еще один пик. На рис. 2.9, Б изображены результаты усиления отбора; в этом случае популяция оказывается сосредоточенной на высшем уровне своего адаптивного пика. Рис. 2.9, В иллюстрирует результаты изменений в среде; в этом случае рельеф ландшафта изменяется, так что адаптивный пик может «рухнуть» под тяжестью популяции, которая таким образом окажется в «долине». Затем под действием отбора популяция начнет подниматься на какой-либо близлежащий пик.
Как показывают рис. 2.9, Л и Б, для того чтобы популяция оставалась на занятом ею пике, необходим непрерывный отбор; такой отбор получил название стабилизирующего отбора: в результате, непрерывного отбора против мутантов популяция сохраняет стабильность в отношении как морфологических, так и физиологических признаков. Процесс В, хотя в его основе лежат те же механизмы, приводит к другим результатам: под его воздействием популяция приобретает новое адаптивное сочетание признаков. Такой отбор получил название направленного отбора.
Рис. 2.9. Несколько адаптивных ландшафтов — вид сверху. А. Ослабление отбора или усиление мутационного процесса. Б. Стабилизирующий отбор. В. Направленный отбор. Г. Случайный дрейф-Д. Популяция, разделившаяся на демы. (Wadd-ington С. Н., An Introduction to Modern Genetics, George Alien a. Unvin Ltd, London, 1939.)
На рис. 2.9, Г представлена небольшая популяция. Здесь случайные события могут вызвать сдвиг в сторону от адаптивного пика, несмотря на отбор, и это та возможность, которую Сьюэлл Райт сформулировал вполне определенно. В принципе каждый индивидуум может случайно стать жертвой несчастного случая или же не найти себе брачного партнера, в результате чего он не внесет своего вклада в следующее поколение независимо от того, сколь хорошо он приспособлен. Поэтому гены, которые он несет, как бы хороши они ни были потенциально, не будут представлены в следующем поколении. Для большой популяции это не будет иметь существенного значения. Однако в небольших популяциях такие случайные события могут оказать серьезное воздействие на частоты генов, так как эти частоты определяются небольшим числом особей. При таких обстоятельствах популяция может медленно отойти от адаптивного пика; процессе этот получил название генетического дрейфа. Подобным же образом при резком сокращении численности популяции или же, что, в сущности, то же самое, при образовании несколькими особями-основателями новой популяции выборка генов из генофонда исходной популяции может произойти неслучайным образом. В результате будет иметь место некое начальное изменение, не связанное с отбором. Это явление известно под названием эффекта основателя.
На рис. 2.9, Д изображена большая популяция, разделенная на ряд субпопуляций, каждая из которых занимает части данного адаптивного ландшафта, несколько отличающиеся друг от друга. Такие популяции, вероятно, встречаются довольно часто. Степень отклонения каждой группы (в результате отбора или дрейфа) от среднего генетического состава популяции в целом зависит от интенсивности обмена генами (то есть миграции индивидуумов) между группами. Для того чтобы противодействовать отбору, поступление данного гена в субпопуляцию должно превышать его элиминацию отбором, а для того, чтобы противодействовать дрейфу, — превышать случайные потери этого гена. Когда такие субпопуляции более или менее изолированы, их называют демами. Демовая структура потенциально важна для эволюционных процессов, и мы обсудим этот вопрос в разд. 3.6. Генетический дрейф иногда называют эффектом Сьюэлла Райта и считают, что его действие направлено против действия отбора (рис. 2.9,Г). Однако, как это понимал Сьюэлл Райт, генетический дрейф может дать популяциям возможность покинуть локальные пики, пересечь имеющиеся в адаптивном ландшафте долины и подпасть под влияние факторов отбора, связанных с новыми пиками. Следовательно, генетический дрейф может способствовать адаптации, но может и препятствовать ей.
2.4. Выводы и резюме.
В процессе исторического развития генетических основ эволюции мы были свидетелями ряда важных переключений внимания с одних факторов на другие.
1. Сторонники Менделя уделяли главное внимание последствиям скрещивания пар особей с известной генетической конституцией. Их основное допущение заключалось в том, что признаки контролируются дискретными факторами, которые не сливаются друг с другом.
2. Холдейн и Фишер рассматривали эволюционное изменение, исходя из частот отдельных генов в популяциях.
3. Для Райта, однако, главными были популяции генов (то есть величины W, а не w), а Добржанский предложил обозначать их термином «генофонд» (см. Th. Dobzhansky, Genetics and the Origin of Species, Columbia Univ, Press, 1937).
4. Начиная с работ Вейсмана, ученые постепенно приближались к открытию физической основы наследственности. В конечном итоге это привело к переосмыслению концепций генов,, мутаций и т. п. в свете представлений о ДНК и синтезе белков.
Первая из этих позиций соответствует менделевской генетике, вторая и третья — популяционной генетике, а четвертая — молекулярной генетике. Позиции 2 — 4 составляют неодарвинизм.
Здесь уместно еще раз привести взгляды, которых придерживается большинство неодарвинистов. (Не все неодарвинисты разделяют в точности одни и те же взгляды, и различия в расстановке акцентов не раз оказывались причиной возникавших & дальнейшем разногласий.) Главные допущения неодарвинизма состоят в следующем.
1. В основе эволюции лежат генетические изменения.
2. Мутации (в их современном общем смысле) представляют собой в конечном счете источник изменчивости. Это могут быть мелкие (точковые) или крупные (хромосомные) мутации, однако мелкие мутации скорее могут привести к благоприятным изменениям, чем крупные, а поэтому они играют более важную роль в эволюции. Принципиально важное значение имеет тот факт, что эти мутации случайны, иными словами, они не направленны.
3. Центральная догма и принцип Вейсмана принимаются.
4. Эволюция осуществляется путем изменения частот генов.
5. Эти изменения могут происходить в результате мутаций, поступления генов в популяцию и оттока их из нее, случайного дрейфа и естественного отбора, причем отбору принадлежит наиболее важная роль.
6. Перечисленных принципов достаточно, для того чтобы объяснить разнообразие и адаптацию организмов на земном шаре.
Осталось еще несколько вопросов, к которым мы вернемся в дальнейших разделах. Это, во-первых, вопрос об • относительном участии генотипа и среды в формировании фенотипа (см. разд 3.2 и гл. 4). Во-вторых, кратковременные изменения генных частот в популяциях мало говорят нам о сохранении самих этих популяций в течение длительного времени. Между тем мы интуитивно ощущаем, что это последнее представляет собой по меньшей мере столь же важную меру эволюционного успеха, как и первое. Можно ли определить это долговременное сохранение с позиций неодарвинизма? И совместимо ли оно вообще с неодарвинизмом? Эти вопросы будут рассмотрены в разд. 3.6 и 5.4.
2.5. Рекомендуемая литература.
Можно назвать ряд хороших сводок по современной генетике: Гейл (7), Айала и Киджер (1), Берри (2). В этих работах используются почти исключительно коэффициенты отбора, тогда как изложение Риклефса (9) основывается На мальтузианском параметре (см. также Кук (4).) Несколько сложнее учебники Раухгардена (10) и Чарлзуорта (3), в которых рассматриваются популяции с перекрывающимися поколениями. Наиболее ясным изложением количественной генетики и, в частности, наследуемости (/i2) все еще остается книга Фальконера (6). С молекулярной генетикой можно познакомиться в книгах Уотсона (12) и Вудса (15). История генетики кратко и интересно изложена Лудовичи (8), а крайне субъективное, но тем не менее интересное описание бурного развития молекулярной генетики принадлежит Уотсону (13). Молекулярная генетика развивается так стремительно, что очень трудно уследить за всеми происходящими в ней событиями. Оригинальное изложение ее развития, дающее определенное представление об увлекательности предмета и о ряде важных его аспектов, можно найти в работе Уотсона и Туза 114).
Глава 3. АДАПТАЦИЯ.
3.1. Введение.
В гл. 2 мы определили приспособленность, исходя из относительного распространения генов в генофондах, то есть исходя из эффекта естественного отбора. Однако связь между распространением данного гена и самим геном зависит от того, как он взаимодействует со своей средой, а это взаимодействие он осуществляет не непосредственно, а через находящегося под генным контролем посредника — через фенотип. Поэтому полезно рассмотреть, почему в данных экологических условиях некоторые фенотипы способствуют передаче генов лучше, чем другие (более приспособлены), иными словами, рассмотреть причину отбора. Подобный подход носит название адаптационистской программы и сочетает в себе рассмотрение формы и функции фенотипов наряду с их экологией и эволюцией.
Со времен Дарвина такая адаптационистская программа всегда пользовалась популярностью у биологов, однако центр тяжести ее постоянно смещался. Как указывал, например, Доукинс (12, 13), дарвинисты пытались рассматривать признаки с точки зрения их полезности для выживания и размножения отдельных особей (то, что Доукинс называет подходом с позиций эгоистичного организма), тогда как после неодарвинистского переворота главное внимание стали уделять объяснению признаков с точки зрения их полезности для переноса генетических единиц (то есть генов и сцепленных групп генов). Этот новый взгляд на вещи привел к тому, что Доукинс называет подходом с позиций эгоистичного гена. При этом фенотипические признаки должно оценивать лишь с учетом того, какое влияние они могут оказывать на распространение тех генов, которые их контролируют. Наконец, признаки организма иногда оценивают в свете того, какую пользу они могут принести той группе (например, семье, популяции, виду), в которую входит данный организм. Такой подход можно назвать подходом с позиций неэгоистичного организма.
Большинство активных приверженцев адаптационизма придерживаются подхода с позиций эгоистичного организма, поскольку, по их мнению, такой подход более или менее совместим с концепцией эгоистичного гена, то есть рассматривает фенотипические признаки с точки зрения их значения для выживания и размножения организмов — носителей этих признаков, что в свою очередь должно быть связано положительной корреляцией с распространением генов, кодирующих эти фенотипические признаки. Дальнейшие объяснения этой методологии и соответствующие примеры будут приведены в разд. 3.2—3.5. Подход с позиций неэгоистичного организма рассматривается в разд. 3.6. Проблемы, в изучении которых были использованы оба подхода— с позиций эгоистичного и неэгоистичного организма,— обсуждаются в разд. 3.7; и, наконец, в разд. 3.8 мы вернемся к разбору взаимоотношений между эгоистичным геном и эгоистичным организмом.
3.2. Методология.
Существуют два различных, но, как станет очевидным, взаимозависимых пути, по которым развивалась адаптационистская программа (5).
Метод 1 (апостериорный!сравнительный подход). При этом методе изучают и сравнивают признаки у близких видов в разные периоды истории данной группы (то есть по ископаемым остаткам) или же — в современной фауне — у видов, занимающих различные экологические ниши. Затем (после такого изучения) дается объяснение обнаруженных различий в адаптивном плане. В отношении различий, коррелированных со временем, принимается допущение, что изменения в признаках следуют за изменениями экологических условий и (или) представляют собой прогрессивные адаптивные усовершенствования. Так, например, направления в эволюции лошадей (рис. 3.1) объясняют переходом их от жизни в лесах к жизни на равнинах. В процессе эволюции наблюдается: 1) увеличение общих размеров, в частности длины конечностей, которые при этом становятся и более стройными; 2) уменьшение числа пальцев до одного и разрастание «ногтя» этого оставшегося пальца с образованием копыта; 3) увеличение резцов в ширину и коренных зубов в высоту вместе с развитием сложных складок твердого цемента на их жевательной поверхности. Изменения 1 и 2 можно рассматривать как адаптации к быстрому передвижению по твердой почве, а изменение 3 — как адаптацию к питанию жесткой степной травой.
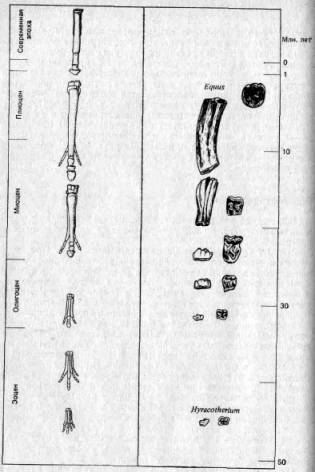
Рис. 3.1. Направления эволюции конечностей и коренных зубов у лошадей (объяснения см. в тексте).
Что касается изменений, коррелированных с пространством, то считается, что дивергенция признаков у популяций, принадлежащих к одному и тому же или к близким видам, произошла в результате различных давлений отбора, испытываемых этими популяциями в тех различных экологических условиях, в которых они находятся. Следовательно, нам важно иметь возможность точно определять различия в тех экологических условиях, под действием которых находятся организмы! Физиологическим экологам, изучающим прибрежную фауну, известно множество примеров близкородственных видов, обитающих в разных частях приливно-отливной зоны и поэтому в разной степени подверженных обсыханию в промежутке между двумя приливами и воздействию волн. Морфологические, физиологические и поведенческие различия между видами, обитающими в разных частях приливно-отливной зоны, во многих случаях можно объяснить как адаптации к этим экологическим различиям; один из таких примеров приведен в табл. 3.1. В этом случае большую устойчивость к высоким температурам и иссушающим условиям у Patella vulgata, обитающей в верхней части этой зоны, чем у P. aspera, обитающей в ее нижней части, можно рассматривать как приспособление к более частому и более длительному пребыванию в воздушной среде в промежутке между приливами. Кроме того, более низкую скорость потери воды в воздушной среде у P. vulgata по сравнению с P. aspera можно отнести за счет различий в форме раковины у этих двух видов (у P. vulgata поверхность, обсыхающая в воздушной среде, меньше); таким образом, морфологические различия поддаются объяснению с позиций адаптационизма.
Таблица 3.1. Некоторые аспекты физиологической экологии морских видов блюдечка (по данным Девиса (10,11))
| Признак | patella vulgata | Patella aspera |
| Зона обитания Выживаемость при t<30°C Скорость потери воды тканями Способность противостоять потере воды | Верхняя Высокая Низкая Высокая | Нижняя Низкая Высокая Низкая |
| Форма раковины | | |
В связи с методом 1 возникает ряд проблем.
1. Не во всех случаях можно твердо установить, являются ли наблюдаемые различия генотипическими или лишь фенотипиче-скими. Так, например, различия в форме раковины, приведенные в табл. 3.1, можно отнести за счет прямого воздействия волн, то есть в этом случае, если бы оба вида развивались в одних и тех же условиях, никаких различий между ними не было бы. Подобного рода негенетические различия (называемые иногда фенотипической пластичностью) в данном контексте интереса не представляют. Отметим, однако, что фенотипическая пластичность может быть детерминирована генетически, а может быть адаптивной. Примеры исследований, в которых был проведен более точный генетический анализ, будут даны в разд. 3.3.
2. Наличие корреляции само по себе нельзя считать доказательством существования причинно-следственной зависимости. Возможно, что в возникновении различий участвуют какие-то скрытые переменные, играющие более важную роль. Возможна также, что различия в форме раковины у блюдечка обусловлены различиями в выеданий хищниками, а это последнее регулируется воздействием волн или обсыханием в промежутке между приливами. В таком случае различия в признаках следовало бы объяснять в свете того, в какой степени раковины определенной формы обеспечивают защиту от хищников, а не от обсыхания или воздействия волн.
3. Не все различия обязательно должны быть адаптивными. Некоторые из них могли возникнуть случайно (дрейф, эффект основателя). Другие могли возникнуть не в результате прямого .отбора, а как «побочные эффекты» признаков, обладающих селективным преимуществом, например в случаях плейотропии (один ген имеет несколько фенотипических эффектов, причем не все они должны быть адаптивными) или аллометрии (см. разд. 4.2). Одним из таких признаков может быть старение. Его нельзя назвать явно адаптивным, поскольку оно, по определению, снижает жизнеспособность и повышает уязвимость затронутых им организмов. Старение может быть адаптивным применительно к группе в целом, например, в том смысле, что оно освобождает место для новых мутаций, связанных с молодыми особями, которые замещают старых особей и, следовательно, стимулируют эволюцию и долговременное выживание группы, однако такую точку зрения с позиций группового отбора разделить трудно (см. разд. 3.6).
В отличие от этого несколько других теорий связывают старение с плейотропными эффектами. Суть их сводится к следующему: в природе организмы гибнут по большей части не от старости, а от несчастных случаев, болезней и хищников. В результате вмешательства этих внешних факторов вероятность выживания часто с возрастом снижается, так что существует некий возраст, после которого животные обычно гибнут и не вносят никакого репродуктивного вклада в популяцию. Если один ген обладает несколькими эффектами, среди которых есть как благоприятные, так и неблагоприятные, то отбор будет направлен на то, чтобы задержать, проявление последних до того периода жизненного цикла, до которого организмы обычно не доживают. Проявление этих гибельных эффектов (внутренне присущие организму факторы смертности) становится заметным лишь тогда, когда внешние факторы смертности устранены, как в лаборатории или в человеческом обществе.- С этой точки зрения старение — не какая-то особая адаптация, а «побочный эффект» генов, которые обладают независимыми от старения благоприятными эффектами, проявляющимися на более ранних стадиях жизненного цикла.
4. Признаки могут быть адаптивными и могут быть обусловлены отбором, но тем не менее различия между ними не обязательно должны иметь селективную основу. Иными словами, для решения одних и тех же проблем могут развиваться различные адаптации, то есть в адаптивном ландшафте имеются многочисленные адаптивные пики, и какой из них займет популяция, зависит от ее начального состава. Это может лежать в основе различия между индийским носорогом, имеющим один рог, и африканским носорогом, у которого два рога. Рога — приспособление для защиты от хищников, и вряд ли можно думать, что один рог — специфическое приспособление для индийских условий, а два — для африканских. Более вероятно, что изначально индийская и африканская популяции несколько различались, в частности по своим морфогенетическим системам, а поэтому под действием одних и тех же селективных сил они достигли различных адаптивных пиков (25).
Метод 2 (априорный/предсказательный подход). Этот метод используют не для объяснения корреляций между фенотипичес-кими признаками и экологической изменчивостью, а для того, чтобы предсказать, какими должны быть эти корреляции. В принципе он позволяет делать предсказания до проведения наблюдений. Подобного рода априорные эволюционные рассуждения часто бывают основаны на допущении, что естественный отбор — процесс оптимизирующий, то есть что в некотором смысле он ведет к эволюции наилучших из возможных признаков. Инженеры и экономисты, которые также стремятся избрать наилучшие решения для конкретных технических или экономических проблем, разработали специальные методы, например теорию оптимального управления, и эти методы находят также применение и в биологии (34). При использовании принципа оптимальности для решения любой проблемы должны быть выполнены следующие основные требования: 1) все возможные решения данной задачи должны быть известны; 2) каждому решению должно быть возможно приписать некоторые числа или сложные математические функции, соответствующие либо стоимости (и), либо цене (с) этого решения относительно какого-либо заранее заданного условия. Математическая задача принципа оптимальности состоит в том, чтобы среди значений v и с найти максимальное v или минимальное с.
Основное условие применимости принципа оптимальности состоит в том, чтобы прибыль и цену можно было измерить, а это зависит в свою очередь от ясного недвусмысленного определения того, какие требования предъявляются к системе. У инженеров и экономистов всегда есть заранее установленные критерии, с которыми можно сопоставлять поведение изучаемой системы. Согласно теории неодарвинизма (см. разд. 3.1), фенотипические особенности биологических систем должны быть такими, чтобы максимизировать расселение потомков (несущих данный признак), генов, а быть может, даже генных комплексов, так что прибыль и цену можно оценивать относительно этих требований. Если бы мы могли точно определять признаки в зависимости от их воздействия на выживание, время генерации и репродуктивный вклад, то было бы относительно легко выбрать те из них, которые максимизируют неодарвинистскую приспособленность. К сожалению, непосредственно вычислить эту величину обычно бывает возможно только для нескольких признаков, и приходится довольствоваться лишь косвенными допущениями. Поэтому весь метод сводится к следующему: 1) допускаем, что отбор максимизирует неодарвинистскую приспособленность (основная гипотеза); 2) переводим 1 в фенотипическую меру приспособленности (вспомогательная гипотеза); 3) используя соответствующие математические методы, находим признак, который максимизирует 2 (или минимизирует ее снижение); 4) сравниваем это предсказание с тем, что наблюдается в природе или обнаруживается в специально созданных экспериментальных условиях. В этой программе адаптационисты редко пытаются опровергнуть основную гипотезу. Они обычно исходят из допущения, что эта гипотеза более или менее верна, а затем пытаются уточнить свое понимание эволюции фенотипа, критически оценивая вспомогательные гипотезы.
Кон (7, 8), а позднее Милсум и Роберж (31) изучали адаптацию применительно к кровеносной системе позвоночных. Они хотели выяснить, какой ширине сосудов будет благоприятствовать естественный отбор. Конечно, оценить диаметр кровеносных сосудов исходя из его воздействия на выживание, скорость развития и размножения не представляется возможным, а поэтому необходима была какая-то вспомогательная гипотеза; такой гипотезой послужило допущение, что отбор должен благоприятствовать самой экономичной структуре. Оно было основано на том, что, чем меньше ресурсов будет затрачено на построение и поддержание кровеносной системы, тем больше их останется у организма для борьбы с болезнями и хищниками и для того, чтобы производить потомство. Кон и его соавторы рассматривали две оценочные функции (рис. 3.2): одна из них возрастает с уменьшением диаметра сосудов (потому что сопротивление трения потоку жидкости при этом возрастает и делает необходимым большую затрату энергии сердцем на прокачивание крови), а другая возрастает с увеличением диаметра (потому что на построение и поддержание системы больших размеров необходимо больше ресурсов). Вспомогательная гипотеза требует, чтобы отбор благоприятствовал той системе, которая обходится дешевле; именно это решение может быть получено графически (рис. 3.2) или из уравнений, определяющих оценочные функции.
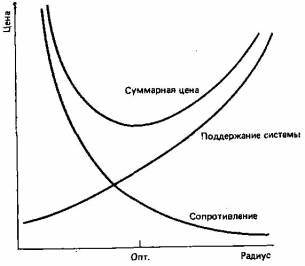
Рис. 3.2. Модель Кона, предсказывающая оптимальный радиус (Опт.) для кровеносных сосудов. Существуют две оценочные функции, одна из которых возрастает, а другая уменьшается с увеличением радиуса сосуда (см. текст). Оптимальным является тот радиус, который минимизирует сумму значений этих функций, то есть представляет собой наиболее экономичное решение.
Подставив в эти уравнения реальные числа, можно вычислить конкретные оптимальные диаметры и сравнить их с результатами наблюдений (табл. 3.2). Кон и его соавторы получили достаточно хорошее совпадение между реальными и ожидаемыми диаметрами. Следует, однако, помнить, что своей работой они не пытались опровергнуть неодарвинизм; они лишь хотели понять, что он означает применительно к развитию кровеносной системы. Данная вспомогательная гипотеза позволяет сформулировать некое онтогенетическое правило, которое может оказаться применимым и для других систем, а именно: системы органов должны быть возможно более экономичными по форме и функции.
Таблица 3.2. Предсказания Кона и результаты измерений реальных сосудов.
| Предсказания | Результаты измерений | |
| Радиус аорты, см | 0,43 | 0,5 |
| Диаметр капилляров, мкм | 2,2 | 4,0 |
Метод 2 также сопряжен с некоторыми затруднениями. Главные из них относятся к формулировке ряда допущенных решений, из которых можно было бы выбрать оптимальное. С одной стороны, мы не можем сделать этого, не прибегая к реальным наблюдениям, то есть метод 2 строится на основе метода 1, а это означает, что модели создаются в соответствии с фактическими данными, а не предсказывают их. С другой стороны, не все мыслимые решения реально осуществимы, а поэтому необходимо некоторым образом ограничить их. Иными словами, оптимизационные методы можно рассматривать как поиски на адаптивном ландшафте (см. разд. 2.3.3) самых высоких пиков, в условиях, когда доступны не все пики: некоторые пики могут соответствовать таким фенотипам, возникновение которых невозможно по морфогенетическим причинам, другие могут оказаться исключенными вследствие исходного генетического состава популяции до начала действия отбора. Возможно также, что уровень генетической изменчивости в популяции окажется недостаточным для достижения наивысшего пика. К сожалению, установить a priori эти ограничения чрезвычайно трудно, и применение метода 2 может дать о них столько же информации, сколько и о самой адаптации.
3.3. Меланизм у бабочек — подход a posteriori.
По мере развития промышленности в Великобритании энтомологи стали замечать, что у многих различных видов бабочек светлая окраска заменяется темной или черной. Особенно ярким примером этого служит березовая пяденица (Biston betularia), темная форма которой получила название carbonaria. Были обнаружены следующие факты: 1) самые ранние находки темных форм были сделаны вблизи районов с сильно развитой промышленностью; 2) самая высокая частота темных форм обнаружена вблизи промышленных центров (отсюда и название этого явления — индустриальный меланизм); 3) меланисты обычно встречаются у видов, которые активны ночью. Апостериорное эволюционное объяснение состоит в том, что светлые формы более заметны для хищников на закопченных ветвях и стволах деревьев, на которых они отдыхают днем. Существуют, конечно, и другие возможные объяснения. Прежде всего меланизм может иметь фенотипическую природу: например, он мог бы возникнуть в результате попадания в организм насекомых промышленных загрязнений. Однако в экспериментах по скрещиванию было показано, что меланизм контролируется генетически и что расщепление по этому признаку происходит так, как если бы он определялся одним доминантным аллелем. Есть и другие возможные объяснения. Так, например, темная окраска могла бы служить защитой от прямого действия загрязнения. Для дальнейшего изучения меланизма у березовой пяденицы Кеттлуэлл (21, 22) провел следующие полевые эксперименты.
1. Чтобы определить, не обладает ли форма carbonaria болы шей приспособленностью, чем типичная березовая пяденица, Кеттлуэлл выпускал меченых особей той и другой формы в промышленных и сельских районах. При повторном отлове в промышленных районах среди выловленных меченых особей было больше carbonaria, а в сельских — больше типичной формы.
2. Равное число особей carbonaria и типичной формы помещали на стволы деревьев в задымленной и незадымленной местностях и непосредственно наблюдали за тем, как их выедали хищники. Оказалось, что в сельской местности хищники уничтожили больше меланистических форм, а в промышленном районе— больше типичных особей.
3. Одна птица — пищуха — поедала без разбору обе формы. Кеттлуэлл указывает, что она кормится, перемещаясь вверх и вниз по стволам деревьев, и поэтому видит силуэт бабочки, сидящей на коре дерева, а не цвет ее крыльев.
Результаты экспериментов 2 и 3 служат особенно убедительными доводами в пользу того, что меланизм возник в ответ на давление со стороны хищников.
Приведенный здесь анализ индустриального меланизма основан на описанном выше методе 1, однако благодаря тщательно проведенным наблюдениям и экспериментам он зашел значительно дальше, чем это обычно бывает или чем это возможно. Кроме того, в этом случае удалось выявить генетическую основу наблюдаемого изменения. Подобного рода программы, основанные на методе 1, но учитывающие также экологические и генетические наблюдения, часто называют экогенетическими — подход, впервые примененный Е. Б. Фордом .(см. (16)) и Добр-Жанским (14).
3.4. Примеры априорного подхода.
3.4.1. Применение принципа оптимальности к адаптациям жизненного цикла
Большая часть рассматривавшихся до сих пор адаптации касалась либо морфологических, либо физиологических признаков. Между тем такие признаки, как характер морфогенетических процессов, скорости роста, общие размеры, продолжительность жизни и вклад в размножение, также, вероятно, подвергались естественному отбору. Все эти признаки участвуют в том, чтобы довести продукты размножения до такого состояния, в котором они могут воспроизводиться, и их называют признаками жизненного цикла или цикла развития. Со времени первой работы Лей-монта Коула (9) об эволюции жизненного цикла писали много (хорошие обзоры принадлежат Стирнсу (37, 38)).
Здесь в качестве иллюстрации мы рассмотрим количество ресурсов, затрачиваемых родительской особью на продуцирование гамет, и продолжительность пострепродуктивного периода ее жизни. То обстоятельство, что не все организмы бурно размножаются один раз в жизни, заставляет думать, что размножение сопряжено с большими затратами ресурсов. В противном случае приспособленность в неодарвинистском смысле была бы всегда максимизирована путем максимально возможного репродуктивного выхода, а следовательно, путем затраты максимального количества ресурсов на образование гамет. Принято считать, что, чем больше ресурсов данная родительская особь вкладывает в размножение, тем меньше их у нее остается для того, чтобы обеспечить защиту собственного организма от несчастных случаев, болезней и хищников, так что эти расходы можно оценивать в терминах выживаемости родительских особей. В результате с увеличением вклада в репродукцию шансы родителей на выживание в пострепродуктивный период снижаются; здесь мы используем фишеровскую меру приспособленности, то есть г (см. разд. 2.3.2), в сочетании с упрощенным вариантом принципа оптимальности, с тем чтобы исследовать, какого рода компромисс возможен между этими переменными.
Значение приспособленности г можно записать как
где It — выживаемость с момента рождения до времени t, a nt — число потомков, произведенных ко времени t (простой вывод этого уравнения см. у Уилсона и Боссерта (45)). В простейшем случае, когда размножение происходит один раз в год (t=1), плодовитость (п) зависит от возраста и родительские особи выживают после размножения, уравнение 3.1 принимает вид
где 1а и // — соответственно выживаемость взрослых особей и потомков, а п— число потомков на одну родительскую особь. Отсюда
Таким образом, из уравнения 6.6 мы видим, что если построить график зависимости 1а от п, то изоклины равных г будут иметь вид прямых с наклоном, равным — l1 (рис. 3.3.).
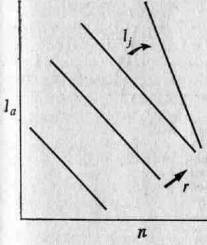
Рис. 3.3. На графике зависимости 1а от п изоклины т представляют собой прямые линии с отрицательными наклонами //. По мере увеличения выживаемости потомков наклон возрастает. В принципе приспособленность, то есть г, возрастает с увеличением la и п.
Однако вследствие компромисса между выживаемостью родительских особей и размножением не все комбинации 1а с п реально осуществимы. Можно представить себе некоторое число распределений реально возможных величин; одно из них показано на рис. 3.4. Здесь выживаемость взрослых особей снижается все быстрее, по мере того как все больше ресурсов вкладывается в размножение, то есть производится все больше гамет.
С помощью этих распределений нетрудно найти оптимальное решение: это та реально осуществимая комбинация 1а и п, которая лежит на самой высокой r-изоклине, то есть максимизирует г, или приспособленность. Для «компромиссной кривой», изображенной на рис. 3.4, этот оптимум приходится на крайнюю правую часть кривой; в этом случае отбор благоприятствует высокому уровню репродукции за счет родительской особи. Такой тип размножения называют семелопарией — родительская особь дает потомство один раз, после чего гибнет подобно лососю.
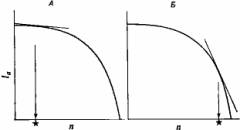
Рис. 3.4. Возможная форма истинной зависимости между 1а и п. А. Выживаемость потомков (//) низкая, "и поэтому отбор благоприятствует небольшому п (оптимум помечен звездочкой) при продолжительном выживании взрослых особей (высокое 1а). Значение // высокое, и поэтому отбор благоприятствует высокому п при низком 1а. Ситуация А ведет к итеропарии, а ситуация Б — к семелопарии (см. текст).
При альтернативном типе размножения, когда достигается некий компромисс между п и 1а, отбор благоприятствует многократному размножению, или итеропарии.
Перейдем теперь к предсказанию. С уменьшением шансов потомков на выживание (то есть с уменьшением /у) наклон г-изо-клин уменьшается и отбор благоприятствует итеропарии. При возрастании // справедливо обратное. (Точный результат зависит от формы «компромиссной» кривой, однако мы продолжаем допускать, что применима только кривая, изображенная на рис. 3.4.) Для того чтобы проверить это предсказание, нам необходимо найти популяции либо того же самого, либо других, но близкородственных видов с различными значениями //. На самом деле точных данных о возрастной специфичности выживаемости для популяций, находящихся в природных условиях, удивительно мало, однако имеются некоторые более или менее эпизодические данные, в общем подтверждающие эти предсказания. Например, пресноводные планарии (трехветвистокишечные тур-беллярии), вылупляясь из яиц, в природе попадают в плохие трофические условия, и, как показали наблюдения (6), молодые особи семелопарических видов гибнут не так быстро, как молодь итеропарических видов, голодающих в лабораторных условиях. Блауэр (3) описал семелопарические и итеропарические виды многоножек фауны Британских островов: число потомков на одну родительскую особь у первых было больше, чем у вторых. Семелопарические виды питаются листьями, которые равномерно покрывают землю в лесу, тогда как итеропарические виды более специализированы, и по крайней мере один из них питается и откладывает яйца на поваленных деревьях, которые разбросаны по лесу неравномерно. Расселение молоди по этим пятнисто распределенным ресурсам носит случайный характер, а поэтому значение // у этого вида ниже, чем у видов, питающихся листьями. Вероятно, по тем же причинам итеропарические виды с непрерывным или полунепрерывным размножением возникли у эндопаразитов, но в этом случае, поскольку родительская особь окружена сверхобильными пищевыми ресурсами — в виде тканей своего хозяина, — она может производить огромное число потомков и не должна расплачиваться за это сокращением продолжительности жизни, о котором говорилось выше.
3.4.2. Приложение теории игр к поведению животных.
Другого рода ограничение, налагаемое на действия особей, направленные на то, чтобы максимизировать приспособленность, связано с активностью других особей, принадлежащих к той же популяции. Можно представить себе, что организмы или по крайней мере признаки, которыми они обладают, как бы играют друг с другом в игры, причем ставка в этих играх—само существование организмов; определить оптимальные стратегии в этих играх может помочь теория игр — один из разделов теории оптимального управления. Используя такой подход, Мэйнард Смит (26) дал определение эволюционно стабильных стратегий (ЭСС) как тех стратегий в эволюционной игре, которые исключают возможность поражения, то есть признаки или сочетания признаков, которые не могут быть вытеснены каким-либо отдельным мутантом. Теория ЭСС оказалась очень полезной для анализа поведения животных.
Наиболее наглядное представление о «поведенческой игре» можно получить, рассмотрев агрессивные взаимодействия между игроками. Возьмем в качестве примера такую игру, в которой возможны только две стратегии (генетически детерминированные наборы признаков, от которых зависит определенный тип поведения, то есть тактика в поведенческой игре) — стратегия ястреба и стратегия голубя. Ястребы всегда дерутся, стремясь поранить и убить своих противников, даже если сами рискуют получить при этом серьезные повреждения, а голуби только разыгрывают агрессивное поведение и никогда не вступают в драку. Какая же стратегия окажется оптимальной? Этот вопрос был досконально изучен Мэйнардом Смитом (27). Допустим, что мы можем численно оценить воздействие данной стратегии на приспособленность и примем для определенности, что эта оценка равна +50 для победителя и 0 для побежденного. Пусть цена времени, затраченного на демонстрацию агрессивного поведения, равна —10, а цена полученного повреждения —100. Если ястреб встречается с ястребом, то в одной половине случаев он, вероятно, окажется победителем, а в другой — будет побежден и получит повреждения, так что в целом плата составит 0,5(50) + + 0,5(—100) =—25. При встрече ястреба с голубем ястреб всегда побеждает, и его плата равна +50; голубь проигрывает, т. е его плата равна 0. Когда голубь встречается с голубем, то оба они демонстрируют агрессивное поведение и каждый из них побеждает в одной половине случаев и оказывается побежденным:—в другой/ Следовательно, плата составит при этом 0,5(50—10)+0,5(—10) == + 15. Эти вычисления можно суммировать в виде следующей матрицы плат (в которой представлены средние платы на одного нападающего):
| Нападающий / Противник | Ястреб | Голубь |
| Ястреб | —25 | +50 |
| Голубь | 0 | +15 |
Какого рода система поведения возникнет в процессе эволюции? В популяции, состоящей из одних голубей (средняя плата — + 15), любые вторгающиеся в нее мутанты-ястребы будут действовать весьма успешно: их плата при каждой встрече с голубем будет равна +50. Следовательно, стратегия голубя не может быть ЭСС. Ястребы распространятся и возьмут верх в популяции, однако при этом средняя плата от каждой встречи превратится в —25 и мутант-голубь окажется в весьма благоприятном положении, поскольку, хотя при каждой встрече с ястребом плата будет равна 0, это все же лучше, чем —25. Следовательно, стратегия ястреба тоже не является ЭСС.
Интуитивно представляется, что стабильной должна быть смешанная стратегия. Пусть Я— доля ястребов в популяции, а 1 —Н — доля голубей. Средняя плата (Ря) для ястреба равна плате для встречи каждого типа, умноженной на
вероятность встретить каждого из противников:
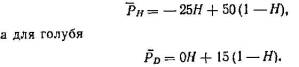
Из этого уравнения можно найти Я, приняв Р0 = Рн (что соответствует ЭСС); отсюда Я=7/12, а 1—Я=5/12. Это стабильное состояние может быть достигнуто при условии, что: 1) каждая особь в данной популяции неизменно придерживается либо стратегии ястреба, либо стратегии голубя, а популяция состоит на 7/12 из ястребов и на 5/12 из голубей; 2) каждая особь выступает в роли как ястреба, так и голубя (смешанная стратегия) с вероятностью 7/12 в первой и с вероятностью 5/12 во второй роли при каждом конфликте.
Игра «ястреб—голубь» иллюстрирует применение теории ЭСС на примере простой и возможно нереалистичной ситуации. Так, по-видимому, лишь в редких случаях «поведенческие игры» состоят всего из двух резко различающихся тактик. Кроме того, приписывать определенное число очков отдельным исходам игры, хотя в принципе это и не сложно, крайне трудно на практике и нередко бывает связано с необходимостью создания сложных вспомогательных гипотез. Тем не менее теория ЭСС широко используется в экологии поведения и оказывает значительную помощь в изучении сложных «поведенческих игр» (23). Мы вернемся к ней в разд. 3.7.
3.5. Изменчивость в природных популяциях; ее значение для адаптации.
Изменчивость вносится в популяцию в результате мутационного процесса (или иммиграции). С возникновением изменчивости общий набор генотипов может значительно увеличиться в результате рекомбинации (некоторые авторы считают это главным преимуществом полового размножения по сравнению с бесполым; см. разд. 3.7), но эта генетическая изменчивость полностью ограничена изменчивостью аллелей, которые могут участвовать в рекомбинации. Дарвин считал, что естественный отбор ограничивает эту изменчивость до уровня адаптивного подмножества, а основная теорема Фишера также подразумевает эволюцию, ведущую от большей изменчивости к меньшей. Исходя из этого, следует ожидать, что популяции эволюционируют в направлении гомозиготного дикого типа и мало отклоняются от этого адаптивного оптимума. Между тем в природных популяциях существует значительная изменчивость, то есть генетический полиморфизм, и экогенетики потратили немало времени и усилий, чтобы собрать соответствующие фактические данные.
Существуют две главные группы доказательств генетического полиморфизма.
1. Прямое наблюдение полиморфизма по признакам, генетическая детерминированность которых известна (например, меланизм у бабочек, характер полос и цвет раковины у улиток, хромосомные аберрации у дрозофилы, группы крови у человека). 2. Данные о структуре ферментов, полученные методом электрофореза. Замещения отдельных аминокислот в молекуле фермента могут изменить ее суммарный заряд в достаточной степени, чтобы изменить скорость перемещения молекулы в электрическом поле. Это позволяет сравнивать сходные молекулы, выделенные из разных особей, и оценивать их изменчивость.
Поскольку замещения в молекуле аминокислоты отражают замещения в молекуле нуклеиновой кислоты, они могут служить показателем генетической изменчивости. Этот метод и его объяснение иллюстрирует рис. 3.5. В табл. 3.3 приведены данные о полиморфизме и гетерозиготности у ряда видов, полученные методом электрофореза.
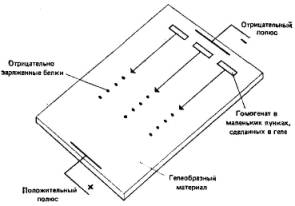
Рис. 3.5. Горизонтальный гель-электрофорез. Пластмассовую пластинку с гелем помещают в сосуд, содержащий соответствующий буфер. Го-могенаты вносят в лунки, сделанные в геле, и пропускают через него электрический ток, под действием которого находящиеся в лунках белки мигрируют на определенные расстояния, зависящие от их аминокислотного состава (см. текст). Затем, используя специфичные красители, можно выявить определенные белки. Одни и те же белки, выделенные из разных животных, могут слегка различаться по аминокислотному составу и поэтому мигрируют на разные расстояния (36).
Таблица 3.3. Доля локусов, в которых наблюдалась изменчивость по ферментам, и доля локусов, в которых была обнаружена гетерозиготность (по данным Шоррокса (36)).
| Вид | Число популяций | Число локусов | Доля полиморфных локусов | Гетерози-готность на 1 локус |
| Человек | 1 | 71 | 0,28 | 0,067 |
| Мышь | 4 | 41 | 0,29 | 0,091 |
| Гольян | 1 | 24 | 0,25 | 0,068 |
| Морская звезда (Nearch-aster aciculosus) | 2 | 24 | 0,71 | 0,213 |
| Мечехвост (Limulus poly phemus) | 4 | 25 | 0,25 | 0,061 |
| Drosophila pseudoobscura | 10 | 24 | 0,43 | 0,128 |
| Drosophila willistoni | 10 | 20 | 0,81 | 0,175 |
Как результаты наблюдений, так и электрофоретические данные позволяют считать, что в популяциях существует значительный полиморфизм и что он, по-видимому, относительно стабилен. Но и классический дарвинизм, и неодарвинизм предсказывают разрушение изменчивости, и в связи с этим между теоретической популяционной генетикой и экогенетикой возникло некоторое напряжение. Существует две возможности справиться с этим парадоксом.
1. Балансовая теория утверждает, что естественный отбор может стабилизировать изменчивость, если существуют гетерозиготность и расщепление генов (Дарвин, придерживавшийся теории слитной наследственности, не мог этого предвидеть). Так называемый уравновешивающий отбор может быть обусловлен: а) преимуществом гетерозигот — гетерозигота, обладающая превосходством (WAA < WAa > Waa), будет, по определению, способствовать сохранению гомозигот; б) отбором, зависящим от частоты; в этом случае приспособленность является функцией частоты генов, так что по мере возрастания частоты данного гена его приспособленность снижается и отбор начинает благоприятствовать другому гену до тех пор, пока частота последнего не возрастет и не произойдет обратное. Такая ситуация может быть создана хищником, всегда выедающим особей с более обычными генотипами. Подобное же действие может оказать выбор брачного партнера; так, например, самки дрозофилы, если им предоставляется возможность выбирать брачного партнера, чаще, по-видимому, спариваются с самцами, обладающими редкими признаками; в) изменениями в давлении отбора в пространстве (различным генотипам благоприятствуют условия в разных частях местообитания, как это показано на рис. 3.6) и во времени (например, циклические изменения метеорологических условий, благоприятствуют одному генотипу в одно время года и другому — в другое).
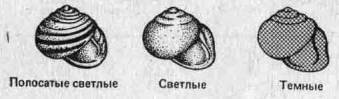
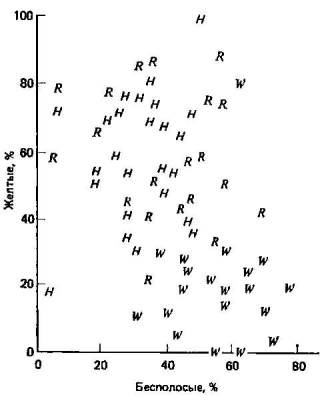
Рис. 3.6. Как известно, полиморфизм по окраске раковины у наземной улитки Cepaea nemoralis вызывается изменением давления отбора в разных местообитаниях. Этих моллюсков поедают дрозды, которые находят их, пользуясь зрением. Светлые полосатые раковины трудно заметить среди низкой растительности (трава R и зеленые изгороди Я). В темном лесу (W) труднее всего увидеть раковины без полос, с однородной темной окраской. (Luria et al., A. View of Life, Benjamin Publ. Co., 1981. Классическое оригинальное исследование полосатости раковины у Сераеа .принадлежит Кейну и Шеппарду (4).)
2. Согласно неоклассической теории, отбор либо имеет место, и тогда он является направляющим, либо он отсутствует, и тогда аллели селективно нейтральны. Предполагается, что полиморфизм по белкам в большинстве случаев "относится к последнему типу. Известно, например, что в молекуле белка лишь относительно небольшая часть имеет существенное значение для его функции, а остальная служит «наполнителем». Замещения аминокислот в этом наполнителе не влияют на функцию белка, но изменяют заряд молекулы.
Разногласия в этих вопросах все еще существуют. Частично проблема заключается в нашей неспособности доказать негативное утверждение: если мы неспособны продемонстрировать наличие отбора, это не означает, что о« отсутствует. Ведь найдено же адаптивное объяснение для изменчивости белков в. .случаях,, в которых, как прежде считалось, она отсутствует. Доказано, например, что изменчивость фермента алкогольдёгидроге-назы у Drosophila melanogaster оказывает влияние на каталитическую активность, устойчивость к нагреванию, субстратную специфичность и удельную активность этого фермента. Поэтому возможность равновесия, обусловленного пространственной: или временной изменчивостью, например, температуры или наличия подходящего субстрата, представляется достаточно реальной. Вместе с тем обнаружение нескольких случаев, связанных с уравновешивающим отбором, нельзя считать доказательством его всеобщности. Для решения этой проблемы необходимо провести подробное исследование на большом числе локусов с учетом как экологических, так и генетических факторов.
3.6. Адаптации на благо группы и адаптации групп.
До сих пор теория неодарвинизма развивалась на основе допущения, что частота того или иного гена возрастает, если он детерминирует признак, который повышает выживаемость и (или) репродукцию его обладателя. Между тем у животных можно найти много примеров такого поведения, которое, очевидно, не способствует выживанию проявляющих его особей. Например, у общественных насекомых некоторые касты лишены способности к размножению, с тем чтобы они могли более эффективно участвовать в различных работах в улье. Рабочая пчела защищает свой улей, убивая вторгшегося в него чужака, даже если погибает при этом сама. Как могло возникнуть в процессе эволюции такое альтруистическое поведение?
Решить этот вопрос удалось после того, как в наших представлениях о приспособленности был достигнут существенный прогресс благодаря работам У. Д. Гамильтона (19). Его рассуждения основаны на том, что родственные особи несут сходные гены, причем сходство между генами тем больше, чем теснее родство. Поэтому если какой-либо признак может повысить выживаемость других особей, обладающих этим же самым признаком, то это может привести к повышению частоты гена, детерминирующего этот признак, даже если данный носитель признака при этом погибнет. Проявления подобного рода альтруизма, вероятно, выше между особями, связанными близким родством, и соответствующий процесс получил название К-отбора (от англ, kin — родство).
Вклад Гамильтона (19) в развитие этой проблемы состоял в том, что он формализовал эти рассуждения и сделал их более строгими. Допустим, что некий альтруистический признак детерминируется одной парой аллелей, А к а, причем аллель А повышает вероятность альтруистического акта у несущей его особи, например делает более вероятным, что эта особь, рискуя собственной жизнью, будет спасать молодых особей, отвлекая на себя внимание хищника. Совершая этот акт, взрослая особь снижает свои шансы на выживание на величину С и повышает соответствующие шансы молодых особей на величину В. Гамильтон показал, что частота в популяции аллеля А по отношению к аллелю а будет возрастать только в том случае, если родство (rl) между донором и реципиентом (выигрывающим от альтруистического акта) достаточно близкое; выражаясь точнее, rl должно быть больше чем С/В.
Например, в случае родители—потомки вероятность того, что любой из генов, имеющихся у одной из родительских особей, окажется у ее потомка, равна 0,5, поскольку потомок наследует гены от обеих родительских особей. Следовательно, rl= = 0,5, и в соответствии с утверждением Гамильтона альтруистический признак сможет спасти более чем двух потомков, прежде чем частота аллеля А повысится относительно частоты аллеля а. В действительности величины rl выражают вероятность того, что организмы, связанные друг с другом определенными родственными отношениями, будут нести одни и те же гены, а тем самым — вероятность того, что реципиент получит гены, определяющие альтруистический признак (несколько других примеров приведено в табл. 3.4). При альтруистических взаимоотношениях между соответствующими родичами с уменьшением rl действия донора должны приносить пользу все большему и большему числу реципиентов; иначе гены альтруизма распространяться не будут. Следовательно, вероятность сотрудничества выше между близкородственными особями.
Идеи Гамильтона помогают понять даже некоторые аспекты организации сообщества у перепончатокрылых. У этих насекомых самки развиваются из оплодотворенных яиц и поэтому
Таблица 3.4. Коэффициенты родства (rl) между особями при разной степени родства.
| Родител и / потомки | 0,5 |
| Бабки-деды /вн уки | 0,25 |
| Полные сибсы | 0,5 |
| Полусибсы | 0,25 |
| Дядя/племянник | 0,125 |
| Двоюродные сибсы | 0,125 |
диплоидны, но самцы развиваются из неоплодотворенных яиц и поэтому гашгоидны. В популяции, в которой особи обоих полов диплоидны, коэффициент родства между сибсами такой же, как между одной из родительских особей и ее потомком (г/ = 0,5; см. табл. 3.4). Однако у самки перепончатокрылых в результате гапло-диплоидии больше общих генов с родной сестрой, чем с собственными дочерьми; то есть генотипы сестер идентичны по всем генам, которые они получают от своих отцов (у него всего один набор), и в среднем по половине генов, которые они получают от своих матерей. Поэтому коэффициент родства между родными сестрами у перепончатокрылых равен не 0,5; а (0,5) X Х(1) + (0,5) (0,5)— 0,75. Таким образом, в этом случае дочерям следует помогать своим матерям охранять яйца и снабжать кормом молодь, а не производить собственных потомков, так что генетическая композиция перепончатокрылых предрасполагает их к развитию такого сообщества, в котором стерильные рабочие самки заботятся о своих родных сибсах. Из всего этого вытекают и дальнейшие следствия, для ознакомления с которыми мы отсылаем читателя к работе Гамильтона (20). Сравнительно недавно идеи Гамильтона были использованы в изучении общественной жизни высших организмов — птиц, млекопитающих и даже человека (см. (44); критические замечания см. (35)).
К-отбор не может автоматически вести к закреплению признаков, полезных для данного вида (то есть обеспечивающих ему длительное существование; см. разд. 2.4); он ведет лишь к признакам, полезным для родичей. Тем не менее можно представить себе способы эволюции таких признаков «на благо вида» путем отбора самих групп, а не особей внутри этих групп. Представим себе две полуизолированные популяции какого-либо вида хищников. В одной из них возникает мутация, порождающая «сверххищника», который обнаруживает и схватывает жертву более эффективно, чем другие особи. Частота такого мутанта в популяции должна возрастать, несмотря на то, что это приведет к вымиранию жертвы, а в конечном счете и самого хищника. В другой популяции такая мутация не возникла, так что она продолжает существовать и в конце концов может захватить пространство, которое занимала вымершая популяция. Аналогичный результат получится в том случае, если от популяции «сверххищников» отделится субпопуляция, не содержащая гена «сверххищника», например, вследствие эффекта основателя.
Можно привести и несколько иной пример: представим себе снова две субпопуляции, но состоящие на этот раз из бессмертных особей. В одной из них появляется мутация, вызывающая старение, и это оказывается выгодным для данной группы, потому что ограничивает ее численность, «очищает» от изношенных особей и увеличивает простор для благоприятных мутаций. Группа, в которой возникло старение, сохраняется дольше другой группы. Этот пример отличается от примера со сверххищниками, потому что он связан с эволюцией признака, положительного для группы, — старения, а не отрицательного — «быть сверххищником». Труднее представить себе, как это может реализоваться. Как, например, ген старения может закрепиться в субпопуляции? Для закрепления такого гена необходимы либо повторные мутации, либо дрейф, либо эффект основателя, поскольку он не может закрепиться при помощи отбора: бессмертные особи оставляют больше потомков, чем смертные, так что ген, обусловливающий старение, будет элиминироваться.
Какие существуют данные за и против группового отбора?
1. Возможен ли групповой отбор? Для того чтобы групповой отбор был реально возможен, скорость вымирания групп и (или) скорость образования новых групп (не имеющих эгоистичного гена или имеющих альтруистический ген) должна быть выше скорости поступления эгоистичных генов в результате либо мутационного процесса, либо, что более важно, обмена генами между подгруппами. По мнению одних биологов, это слишком жесткое требование, чтобы групповой отбор мог быть обычным явлением (например, (24), тогда как другие этого не считают (например, (17, 43)).
2. Происходит ли групповой отбор? Располагаем ли мы данными о существовании признаков, важных для сохранения группы, но нестабильных при индивидуальном отборе? Один из таких возможных признаков — это старение, и оно широко распространено. Однако его можно объяснить также, как плейот-ропный эффект благоприятных генов, которые отбирались обычным образом (см. разд. 3.2). Другой возможный пример — способность популяций регулировать свою численность, поддерживая ее на таком уровне, чтобы не допускать чрезмерного изъятия ресурсов. Винн-Эдвардс в своей знаменитой книге «Расселение животных и его зависимость от социального поведения» (Wynne-Edwards, Animal Dispersion in Relation to Social Behaviour, Oliver a. Boyd, 1962) привел многочисленные примеры такого рода явлений. Однако при более тщательном изучении то, что представляется такой саморегуляцией, на самом деле часто можно объяснить как результат зависящей и не зависящей от плотности регуляции, обусловленной внешними факторами — хищниками, погодой и даже самими ресурсами. Многие другие групповые признаки оказалось возможным объяснить К-отбором.
Данные, свидетельствующие за и против группового отбора, неубедительны, и поэтому данная проблема продолжает вызывать разногласия. Представляется вероятным, что в некоторых системах может происходить и действительно происходит групповой отбор, но вопрос о его относительной частоте по сравнению с индивидуальным отбором остается открытым. Возможно также, что существует групповой отбор сходного типа, действующий, однако, не во внутривидовых группах, а на уровне вида в целом (см. гл. 5).
3.7. Адаптации самого полового размножения как такового и порожденные им адаптации.
Половое размножение большинства высших организмов характеризуется следующими важнейшими чертами: 1) продуцирование гамет в результате мейоза; 2) рекомбинация элементов генома путем кроссинговера (см. разд. 2.1.2); 3) случайное распределение гомологичных хромосом по всем продуктам мейоза (см. разд. 2.1.1); 4) создание новых особей в результате сингамии (слияние двух гамет, обычно от двух отдельных особей). Благодаря этим процессам половое размножение может порождать значительные геномные различия между родительскими особями и их потомками даже в отсутствие мутационного процесса. Этот способ размножения широко распространен, встречается по всему животному миру и, вероятно, возник очень давно.
Существуют также и другие способы размножения, в которых участвуют одна родительская особь и гаметы одного типа. Эти способы известны под общим названием партеногенеза. Известно несколько различных типов партеногенеза. При арре-нотокии самцы развиваются из неоплодотворенных яиц, как у некоторых перепончатокрылых, у которых самцы гаплоидны, а самки диплоидны. При телитокии самки развиваются из неоплодотворенных яиц, которые образуются либо в результате митоза (мейоз подавлен) — процесс, известный под названием апомик-сиса, либо в результате модифицированного мейоза, при котором диплоидность восстанавливается после деления путем слияния гаплоидных ядер (аутомиксис). При апомиксисе единственным источником изменчивости служит мутационный процесс, так что потомки неизменно бывают похожи на родительских особей. При аутомиксисе изменчивость может возникать в результате кроссинговера, однако постмейотическое слияние ядер повышает гомозиготность, а поскольку при этом возрастают шансы на экспрессию вредных генов, которые обычно бывают рецессивны, то он встречается редко. Поэтому мы сосредоточим внимание на апомиктическом партеногенезе, имеющем более широкое распространение. Следует также отметить, что у некоторых видов телитокия сочетается с половым размножением (например, чередование полового и партеногенетического размножения у коловраток и тлей, связанное с различными внешними условиями). Отметим также, что используемая в этой области терминология часто создает путаницу, так как ботаники и зоологи пользуются разными системами терминов. Приведенной выше терминологии придерживаются зоологи; с другими терминами можно ознакомиться у Мэйнарда Смита (28).
Самый обычный и широко распространенный способ размножения—это, безусловно, половое размножение, и все же в смысле продуцирования потомков наиболее эффективен, по-видимому, партеногенез. Рассмотрим, например, двух самок, сходных физиологически, но одна из которых несет мутацию, ведущую к апомиксису. Обе самки продуцируют за счет пищи, которую они съедают, равное число яиц, но из всех яиц партеноге-нетической самки развиваются тоже самки, которые тоже будут размножаться партеногенетически, а среди потомков половой самки самок только половина, а остальные — самцы. В принципе, таким образом, система полового размножения терпит 50% убытка (так называемая плата за пол) по сравнению с парте-ногенетической в смысле продуктивности, а поэтому партеногенез должен распространяться в популяции за счет полового размножения. Следовательно, половое размножение не является ЭСС (см. разд. 3.4.2) и его широкое распространение нуждается в объяснении.
Имеется ряд потенциально возможных объяснений этой загадки, которые были рассмотрены Уильямсом (42), Мэйнардом Смитом (28) и Беллом (2). Их можно грубо разделить на зависящие от группового отбора и не зависящие от него.
1. Объяснения, основанные на «благе для группы». Как уже говорилось, половое размножение порождает разнообразие внутри популяций. Кроме того, в результате расщепления оно создает возможность для отделения благоприятных мутантов от определенных генных комплексов, так что они могут распространиться по всей популяции, а также для отделения благоприятных генных комплексов от неблагоприятных мутантов. Партеногенез, однако, ограничивает разнообразие (при апомик-сисе оно зависит от возникновения мутаций, что происходит редко; см. разд. 2.3.2), препятствует распространению благоприятных мутаций (поскольку мутации заключены в определенных генных комплексах и могут распространяться только в отдельных клонах) и означает, что неблагоприятные мутации, будучи заключены в геномах, будут накапливаться в популяциях, поскольку они появляются в одном клоне за другим. Последний механизм был назван «храповиком Мёллера», который впервые высказал эту идею (32). Таким образом, популяции, размножающиеся половым путем, должны сохраняться дольше чем партено-генетические популяции.
2. Неодарвинистские объяснения. Здесь в центре внимания находятся воздействия полового размножения на гены, которые его кодируют, и на организмы, которые им обладают. Следует ожидать, что потомки половых родителей будут отличаться большим разнообразием, чем потомки партеногенетических родителей. Поэтому, хотя партеногенетические родители могут быть более плодовитыми, чем половые, у них по сравнению с последними меньше шансов процветать и выжить, если среда, в которой они появились на свет, изменчива в пространстве и (или) во времени по своим физико-химическим и (или) биологическим условиям. Таким бразом, эти различия в выживаемости могут сдвинуть равновесие в пользу полового размножения. Объяснения сторонников группового отбора страдают теми же недостатками, что и объяснения, уже обсуждавшиеся в разд. 3.6. В свою очередь сторонникам неодарвинистских объяснений еще надлежит убедительно доказать, что в реальных средах создания изменчивости достаточно для того, чтобы оплатить расходы, связанные с половым размножением. К сожалению, результаты имеющихся наблюдений довольно бессвязны и создают путаницу, а выработать критерии, которые позволили бы сделать решительный выбор между многочисленными конкурирующими гипотезами, пока не удалось (2). Поэтому дискуссия продолжается.
Однако, каким бы ни был исход дискуссии, не вызывает сомнений, что эволюция полового размножения оказала глубокое и далекоидущее воздействие на сами эволюционные процессы. Некоторые из наиболее важных эффектов приведены ниже.
1. Эволюция генетических систем. Существование полового размножения имеет важные последствия для генетических механизмов. Самое главное из них состоит в том, что гены (или группы сцепленных генов) представляют собой более важные единицы отбора, чем целые геномы, так как последние — недолговечные единицы, разбивающиеся на части при мейозе и смешивающиеся при сингамии. Вызывающие это механизмы и такие связанные с ними явления, как диплоидность и доминирование, сами служат объектами отбора, и поэтому для них напрашивается адаптационистское объяснение. Например, дипло-идность и доминирование создают, возможно, некоторую защиту от вторжения гибельных мутаций, а поэтому следует ожидать, что гены, контролирующие соответствующие признаки, будут распространяться в популяциях. В связи с этим Фишер (15) высказал мнение, что существующий в популяции уровень доминирования эволюционировал путем включения генов-модификаторов. Так, модифицирующий аллель М благоприятен, если под его влиянием аллель А, находящийся в другом локусе, становится доминирующим над вредными мутантами (например, А1). Таким образом, между аллелями А и М существует эписта-тическое взаимодействие (см. разд. 2.1.2). Однако при подобной интерпретации возникает ряд затруднений (обзор см. (39)). Например, аллель М будет иметь лишь очень небольшое преимущество, так как он будет оказывать действие только в сочетании с гетерозиготами АЛ1, а это вначале будет редким. Поскольку М, скорее всего, обладает и другими физиологическими эффектами, а не только модифицирует доминирование, эволюция в этом локусе будет определяться главным образом этими другими эффектами. Однако, как мы увидим в дальнейшем (см. разд. 4.3), доминирование не обязательно должно было возникнуть в процессе эволюции специально для того, чтобы подавлять эффекты мутаций, но может быть проявлением некой неспецифичной способности сопротивляться любым изменениям в характере развития независимо от того, вызываются ли они генетическими факторами или факторами среды. Такого рода проблемы возникали постоянно с тех самых пор, как в 30-х годах были впервые опубликованы книги Фишера (15) и Дарлингтона (С. D. Darlington, The Evolution of Genetic Systems, Oliver a. Boyd, 1939). Оказалось, однако, что эти проблемы на редкость плохо поддаются строгому анализу и экспериментальному исследованию. Связи между причиной адаптации и эволюционными эффектами переплетаются здесь так тесно, что их иногда трудно распознать, не говоря уже о том, чтобы попытаться разобраться в тех или других по отдельности. Тем не менее это важная и развивающаяся область, и Белл (2) назвал ее мета-генетикой.
2. Эволюция гамет. Первоначально все продукты мейоза в данной популяции были, вероятно, одного размера (изогамия), как у многих ныне живущих одноклеточных организмов. Однако увеличение размера гамет, вероятно, давало известное преимущество, потому что развивающаяся зигота получала при этом больше ресурсов и ее шансы на выживание возрастали. Вместе с тем мелкие размеры также обладают преимуществом, позволяя из того же количества ресурсов получить больше гамет. При наличии отбора, направленного на продуцирование крупных гамет, немедленно возникает отбор, направленный на то, чтобы мелкие гаметы находили их и сливались с ними, потому что таким образом гены, содержащиеся в маленькой гамете, приобретают возможность воспользоваться ресурсами, запасенными в большой гамете. Можно представить себе также отбор, направленный на создан.ие у крупных гамет способности противостоять оплодотворению мелкими гаметами, с тем чтобы, соединяясь с крупными гаметами, они производили еще более крупные зиготы. Однако доход в смысле повышения выживаемости, который дает увеличение размера зиготы, по-видимому, подчиняется закону тенденции нормы прибыли к уменьшению, то есть по мере увеличения зиготы жизнеспособность возрастает все меньше и меньше. Поэтому гены, определяющие большие размеры гамет, не могут выдержать натиска генов, определяющих малые размеры, которые к тому же ведут себя «нечестно» (пользуются благами, предоставляемыми им их более крупными партнерами, но не платят за это). Вместе с тем мелкие гаметы не могут устоять против инвазии мутантов вследствие низкой выживаемости мелких зигот. Это рассуждение можно сделать строгим (2, 33) и предсказать, что эволюционно стабильной стратегии соответствует анизогамия, то есть мелкие (мужские) и крупные (женские) гаметы. Как правило, и в царстве растений, и в царстве животных наблюдается именно анизогамия, лежащая в основе всех других различий между полами.
3. Эволюция соотношения полов. Теперь, после того как мы показали, как могло возникнуть разделение популяции на самцов (продуценты мелких гамет) и самок (продуценты крупных гамет), можно задать вопрос: каких соотношений между двумя полами следует ожидать в популяциях? Ответ нам известен — обычно соотношение их составляет 1:1, но если продуценты мелких гамет могут производить на единицу ресурсов больше гамет, чем продуценты крупных гамет, то почему соотношение полов не смещено в пользу самок?
Одно из решений этой задачи может быть найдено с позиций ЭСС, и основы его были заложены еще Фишером (15). Допустим, что в некой популяции на каждого самца приходится по 100 самок и что репродуктивная эффективность каждого самца в 100 раз выше ожидаемой репродуктивной эффективности самки. В таком случае родительская особь, все потомки fi которой мужского пола, оставит в 100 раз больше потомков F2, чем особь, все потомки fj которой были бы женского пола. Поэтому сдвиг соотношения в пользу «дочерей» не представляет собой ЭСС; в точности то же самое можно сказать и о сдвиге его в пользу «сыновей». Только в том случае, если соотношение полов равно 1:1, ожидаемая эффективность для «сыновей» и «дочерей» будет одинакова. Следовательно, такое соотношение и соответствует ЭСС.
В сущности, это предполагает, что продуцирование «сыновей» и «дочерей» обходится примерно одинаково. Допустим, например, что продуцирование «сыновей» обходится дороже (скажем, вдвое), потому что они крупнее «дочерей». При соотношении полов 1 : 1 «сын» производит столько же потомков, сколько «дочь», но, поскольку «сыновья» обходятся дороже, это невыгодное предприятие, так как при этом снижается общее число потомков, которое может оставить одна родительская особь. Поэтому родительским особям выгодно затрачивать свои ресурсы на продуцирование «дочерей». В результате возникает известный перекос в пользу самок, однако по мере его возрастания ожидаемый репродуктивный успех «сыновей» повышается. Равновесие достигается в том случае, если родительские особи вкладывают равное количество ресурсов в продуцирование потомков того и другого пола, а не тогда, когда они производят их в равном числе. Поэтому в тех случаях, когда затраты на продуцирование самок и самцов различаются, следует ожидать отклонения от соотношения полов 1:1. Меткаф (30) обнаружил, что у Pollster metrlcus (перепончатокрылые), у которого самки имеют мелкие, а самцы — крупные размеры, соотношение полов сдвинуто в пользу самок, тогда как у близкородственного вида P. variatus самцы и самки имеют одинаковые размеры и соотношение между ними составляет 1:1. Следует напомнить, однако, что у перепончатокрылых самки могут регулировать пол своих потомков (см. разд. 3.6). У большинства видов соотношение полов определяется половыми хромосомами, и, по всей вероятности, механизм мейоза будет удерживать это соотношение на уровне 1 : 1 даже при наличии полового диморфизма по общим размерам. Таким образом, в этом случае генетические сдерживающие механизмы перевешивают другие силы отбора (см., однако, Т. Н. Glutton-Brock, Nature, 298, 11—13, 1982).
4. Половой отбор. Поскольку самцы продуцируют более мелкие, а поэтому более многочисленные гаметы, чем самки, и поскольку соотношение полов в норме составляет 1:1, между самцами часто наблюдается конкуренция за самок. Давление отбора, направленное на повышение способности самцов «добыть» себе самку, очень сильное, так как плата за поражение очень высока. Такого рода отбор известен под названием полового отбора и был впервые описан Чарлзом Дарвином. Он может действовать, благоприятствуя способности одного пола (обычно, но не всегда мужского) непосредственно конкурировать за обладание другим полом и(или) способности одного пола привлекать другой. Примеры можно найти у Кребса и Девиса (23). Эти авторы указывают, что интенсивность полового отбора зависит от степени конкуренции за брачных партнеров, которая в свою очередь зависит от: а) величины вклада обоих видов в создание потомков (то есть чем меньший вклад вносит один пол в потомков, тем сильнее конкуренция между представителями этого пола за брачных партнеров; обычно это означает, что самцы конкурируют за самок, однако не всегда, как показывает пример колюшки); б) отношения самцов к самкам, одновременно доступным для спаривания (если число последних уменьшается, например, вследствие несинхронного размножения, то конкуренция усиливается).
Аспект полового отбора, который особенно заинтересовал Дарвина и который с тех пор оставался в центре внимания,— это эволюция чрезвычайно сложных демонстраций и украшений подобно хвосту павлина. Такие украшения могут оказаться в известной степени неблагоприятными для своих владельцев, так как их образование, поддержание и демонстрация требуют затраты ресурсов, а между тем они возникают в процессе эволюции довольно часто, принимая иногда фантастические формы. Почему это происходит? До сих пор общепринятого или приемлемого объяснения не найдено, если не считать одного, впервые высказанного опять-таки Фишером (15) и, во всяком случае, правдоподобного. Фишер полагает, что украшения могут сохраняться отбором просто потому, что они привлекают самок. Если в данной полигамной популяции большинство самок отдают предпочтение самцам, имеющим определенное украшение, то мутация, детерминирующая отсутствие этого украшения, не сможет закрепиться в популяции, так как мутантные самцы будут неприемлемы в качестве брачных партнеров, даже если у «сыновей», которых они могут произвести, перспективы на выживание лучше, чем у «сыновей», имеющих украшения. Таким образом, после возникновения украшений у самцов и приверженности к ним у самок эволюция этих украшений протекала как самоусиливающийся процесс. Но в таком случае возникает критический вопрос: как возникли эти украшения? На этот вопрос можно ответить по-разному. Один из ответов состоит в том, что первоначально данное украшение было коррелировано с другими признаками и эффектами, которые способствовали продуцированию потомков и выживанию. Поэтому самки, выбиравшие брачных партнеров с украшениями, обеспечивали большую приспособленность собственным генам. Только после того, как украшение стало более выраженным в результате самоусиления, эта корреляция распалась. Конечно, это всего лишь гипотеза, которую трудно проверить, после того как вступил в действие самоусиливающийся половой отбор.
3.8. «Эгоистичная» ДНК и гены в организмах.
На протяжении всей этой главы различие между эгоистичным организмом и эгоистичным геном оставалось неясным, как это обычно и бывает в адаптационистской программе. В чем же это различие? И как велико его значение? Два этих подхода можно грубо сформулировать следующим образом:
1. Согласно теории эгоистичного гена, отдельные фенотипи-ческие (организменные) признаки возникли в процессе эволюции, потому что они помогают генам реплицироваться.
2. Согласно теории эгоистичного организма, гены, которые становятся наиболее обычными в данной популяции, достигают этого, потому что они способствуют выживанию и размножению организмов.
Поскольку гены «используют» организмы для того, чтобы реплицироваться, и поскольку организмы находятся под контролем генов, эти две точки зрения обычно сводятся к альтернативным способам рассмотрения одного и того же явления. Ген, который действовал за счет других генов данного организма, а следовательно, за счет выживания и размножения этого организма как целого, не мог бы распространяться достаточно эффективно. Это подразумевает, что на уровне организма, по всей вероятности, существует отбор на интеграцию и сотрудничество между генами. Вместе с тем теория эгоистичного гена предсказывает, что гены, которые нейтральны в отношении выживания и плодовитости организмов, могут распространяться. Например, наличие больших количеств нефункциональной ДНК (см. разд. 2.2.3) можно было бы объяснить именно таким образом. Доукинс (12) высказал предположение, что эта избыточная (эту ДНК иногда называют «сорной») ДНК подобна «паразиту... или безвредному, но бесполезному пассажиру, «голосующему» на дороге, с тем чтобы пристроиться на машины выживания, созданные другой ДНК». Главный вопрос здесь состоит, однако, в том, насколько нейтральны отдельные гены и их эффекты и насколько они должны быть нейтральными, чтобы, входя в состав организма, избежать проверки со стороны отбора. Является ли, например, стоимость репликации и несения так называемой сорной ДНК недостаточной, для того чтобы оказывать влияние на выживание и плодовитость носителя? Действительно ли она лишена какой-бы то ни было функции или мы просто до сих пор не сумели эту функцию обнаружить? Некоторые из этих вопросов живо обсуждаются в ряде статей, опубликованных в журнале Nature в 1980 г. (см. 284, 601—607; 285, 617—620; 285, 645—648).
Поскольку гены объединены в организмах, нетрудно понять, почему они должны сотрудничать друг с другом; однако теория эгоистичного гена ставит и другой вопрос: почему организмы? Почему, спрашивает Доукинс (13) в заключительной главе своей книги «Расширенные границы фенотипа», гены собираются в ге-номы, а клетки — в многоклеточные тела? Ответить на это в общей форме несложно: гены, входящие в группы, передаются более эффективно, чем гены, которые в них не входят, однако объяснить причины этого более детально не так просто, и адаптационистской программе следует теперь ими заняться. Кроме того, если интеграция, кооперация и коадаптация так важны, то почему они не привели к чему-то большему, чем сцепление генов? Почему, — и здесь мы вновь возвращаемся к разд. 3.7,— механизмы, связанные с половым размножением, которые в такой же степени склонны разрушать благоприятные сочетания генов, как и создавать их, получили возможность сохраниться? Почему, говоря словами Тренера (41), геном не застывает? Это трудные, но тем не менее важные и глубокие вопросы, к которым адаптационистская программа только начинает приступать.
3.9. Рекомендуемая литература.
Критическое рассмотрение адаптационистской программы см. у Гоулда и Левонтина (18). Обоснование этой программы дает Доукинс (13). Использование принципа оптимальности иллюстрирует Александер (1), а теории игр —Мэйнард Смит (29). Приложение адаптационистской программы к физиологической экологии рассматривают Таунсенд и Кейлоу (40), а к экологии поведения— Кребс и Девис (23).
Глава 4. ЭВОЛЮЦИЯ И РАЗВИТИЕ.
4.1. Введение.
При неодарвинистском подходе к эволюционной биологии главное внимание уделяется генетическим основам изменения и постоянства популяций, тогда как адаптационисты сосредоточили внимание на фенотипах. Однако эти два аспекта эволюции разделять не следует, ибо выражение фенотипа частично зависит от генотипа, а распространение генов зависит от того, сколь успешно фенотипы взаимодействуют со своей средой. Эти взаимозависимости иллюстрирует рис. 4.1. Экспрессия генов осуществляется через посредство морфогенетической системы, на которую оказывают также влияние изменяющиеся факторы среды. Уоддингтон (12) назвал изучение этих взаимодействий эпи-генетикой. В гл. 3 мы уже коснулись экологических и реверти-рованных эпигенетических законов, показанных на рис. 4.1.
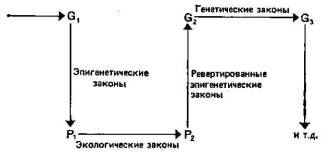
Рис. 4.1. Некое исходное распределение генов (Gi) дает определенные фенотипы (Pi) в соответствии с эпигенетическими законами. Распределение фенотипов изменяется в соответствии с экологическими законами (переходя в pz)- Эти фенотипы вносят в генофонд (Gj) гены, которые распределяются в соответствии с генетическими законами (Оз). В этой главе основное внимание уделено рассмотрению пути от генов (G) к фенотипам (Р).
Здесь мы обратимся к самим эпигенетическим законам. Будут рассмотрены лишь два из наиболее важных принципов: как мелкие мутации могут амплифицироваться в процессе развития и как морфогенетическая система противодействует некоторым типам генетических изменений.
4.2. Амплификация в процессе развития.
Джозеф Нидхем (9) различал в онтогенезе три основных процесса: 1) рост — увеличение объема и веса; 2) развитие — диф-ференцировка и возрастание сложности организации (морфогенез); 3) созревание — прекращение развития и роста и наступление репродуктивных процессов. Джулиан Хаксли (7) изучал, каким образом второй из этих процессов взаимодействует с первым, направляя и регулируя изменения размеров органов и соотнося их друг с другом и с изменениями величины организма в целом. Такие взаимоотношения часто описываются простым математическим уравнением
где У — размеры данной части, X — размеры другой части или организма в целом, а и р — константы (но ср. Смит (11)).
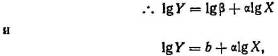
где b=lg(бета). Эти соотношения известны под названием алломет-рических зависимостей, где а — константа аллометрии: если а>1, то У увеличивается в размерах быстрее, чем X, если же <х<1, то наоборот, если а=1, то X и У растут пропорционально одна другой, и этот особый случай называют изометрией. Соответствующие примеры приведены на рис. 4.2. Эти зависимости могут отражать лежащие в их основе морфогенетические ограничения, и (или) скейлинг-эффект, и (или) конкуренцию между отдельными частями организма за ограниченные ресурсы, поступающие с пищей (11).
Нидхем (9) указывал, что как в принципе, так и практически относительные скорости и сроки наступления трех выделенных им фундаментальных процессов можно изменять и что такие регулировки оказывают глубокое влияние на конечный результат развития. Можно, например, экспериментально ускорить дифференцировку по отношению к росту и получить карликов (экспериментальная карликовость). У некоторых беспозвоночных можно подавить размножение и получить великанов (паразиты нередко кастрируют своих хозяев, что приводит к тому же результату). Если возможно искусственно разделить рост и развитие, то можно предполагать, что такая гетерохрония играет важную роль в эволюции. Мелкие мутации могли бы оказывать некоторое влияние на скорости и сроки роста органов и тканей, но сильно воздействовать на конечный результат — иными словами, небольшие изменения в а и (или) b могут иметь очень важные последствия для организации и морфологии взрослого организма.
Рис. 4.2. Примеры аллометрии в теории (Л) и на практике (Б). Построенные в логарифмическом масштабе графики зависимости размеров одного органа (Y) от другого или от размеров организма в целом (X) часто имеют вид прямых. Если наклон этих прямых равен единице, то зависимость называют изометрической (для сравнения на рис. Б она изображена пунктиром); в этом случае X и Y увеличиваются в размерах с одинаковыми скоростями. Зависимость между весом кишечника и общим весом тела у млекопитающих и птиц примерно изометрична. Если наклон больше единицы, то зависимость положительно аллометрична и размеры Y увеличиваются быстрее, чем размеры X (например, данные по крысам). Когда наклон меньше единицы, то зависимость отрицательно аллометрична и Y увеличивается медленнее, чем X (например, данные по собакам). Такие графики можно строить для особей, принадлежащих к одному виду (например, данные по собакам и крысам), или для особей, принадлежащих к нескольким разным видам (например, данные по млекопитающим и птицам).
Олберч и др. (2) систематизировали возможные способы возникновения таких регулировок по отношению к некой временной компоненте, определяющей инициацию развития, возраст и начало созревания (то есть 3-й процесс из триады Нидхема). На рис. 4.3 эта система представлена графически. В каждом квадрате заключена некая траектория развития, то есть некий показатель формы в зависимости от размеров или возраста. (Если используются размеры, то эти графики можно рассматривать как эквиваленты зависимостей, которые исследовал Хаксли.) Сплошными линиями изображены траектории предков, а прерывистыми — потомков. Вертикальная линия «Старт» указывает начало развития, а линия «Стоп» — его прекращение. При всех изменениях в верхнем ряду развитие замедляется или урезывается по сравнению с развитием предков, так что зрелость наступает на более ранней стадии развития. Этот процесс называется педоморфозом. При всех изменениях в нижнем ряду развитие, напротив, ускоряется или удлиняется, и это носит название пераморфоза. В этих случаях потомки проходят через предковые стадии на более раннем этапе развития, то есть имеет место рекапитуляция предковых форм.
Можно привести примеры каждого из этих процессов.
1. Ретардация/неотения. Классическими примерами служат некоторые амфибии (хвостатые), у которых взрослые особи сохраняют жабры и другие личиночные органы. Взрослая форма выглядит поэтому как личинка и обитает в воде.
2. Прогенез. Самцы некоторых ракообразных достигают половой зрелости в то время, когда их общие размеры невелики. Эти миниатюрные самцы «паразитируют» на самках, которые гораздо крупнее (иногда на несколько порядков).
3. Предварение смещения (divdisplacement). У мутантных «ползающих» (screeper) кур кости ног дифференцируются позднее, чем у нормальных цыплят, но дальнейший их рост происходит с обычной скоростью. Время вылупления также не изменяется, а поэтому у вылупляющихся цыплят ноги короткие.
4. Акселерация. У некоторых сравнительно молодых видов аммонитов рост лопастных линий на раковинах ускорен по сравнению с предковыми видами. Однако в этом замешаны и другие процессы, так что в дальнейшем данный пример будет рассмотрен более подробно.
5. Гиперморфоз. Гигантские по сравнению с другими видами размеры ныне вымершего оленя Megaloceros giganteus достигались, возможно, за счет продолжения роста и задержки размножения.
6. Задержка смещения (postdisplacement). Этот процесс вместе с предварением смещения, возможно, участвует в регуляции типов полосатости у зебры. Бард (3) считает, что у зародыша имеется единственный механизм, создающий вертикальные полосы. В зависимости от того, на какой стадии роста он вступает в действие, полосы на голове будут шире (процесс начинается раньше — предварение смещения) или уже (процесс начинается позднее — задержка смещения) и расположение полос может быть весьма разнообразным. Наблюдается несколько типов полосатости, которые можно объяснить инициацией процесса образования полос в течение третьей (Equus burchelli), четвертой (Е. zebra) или пятой (Е. grevyi) недель развития.
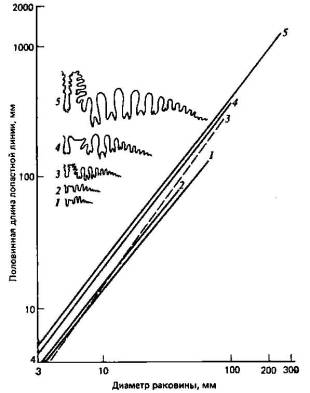
Рис. 4.4. Участие нескольких морфогенетических явлений в эволюции аммонитов (8). Изображены лопастные линии предковых (/) и происходящих от них видов (2—5). Самая древняя часть лопастной линии — небольшой участок справа. Более подробные объяснения см. в тексте.
Не следует, однако, ожидать, что нам всегда удастся наблюдать «в чистом виде» описанные выше изменения; в сущности, более вероятно возникновение морфологических изменений в результате сочетания различных морфогенетических явлений. Хорошим примером служит аллометрическая зависимость между длиной лопастной линии и общими размерами тела у аммонитов, о которой уже говорилось выше и которая представлена на рис. 4.4. Здесь наклон кривых для потомков круче, чем для предков (рис. 4.4,1), так что дифференцировка этих лопастных линий у более молодых видов ускорена по сравнению с предковыми видами. Но, кроме того, в филогенезе этой группы наблюдается выраженное увеличение общих размеров, и у потомков продолжается аллометрический рост лопастных линий, выходящий далеко за пределы размеров взрослых особей у предковых видов (гиперморфоз). Наконец, траектории видов-потомков располагаются выше, чем траектории предковых видов, так что потомки с самого начала уже обладают известным преимуществом, возможно, в результате какого-то предварения смещения.
Итак, все приведенные выше примеры показывают, каким образом изменения процессов развития могут оказывать существенное воздействие на дефинитивную морфологию взрослого организма. В принципе это может быть достигнуто при помощи мелких изменений в скоростях нескольких ключевых процессов, то есть путем мелких генетических изменений. В этом и заключается амплификация в процессе развития.
4.3. Канализация развития.
Одно из важнейших заключений, вытекающих из эмбриологических исследований, состоит в том, что развитие — процесс упорядоченный, способный в значительной степени сопротивляться нарушающим воздействиям, оказываемым на него «извне» (например, экспериментальные воздействия) или «изнутри» (например, мутации). Это можно проиллюстрировать следующими примерами.
1. Специалисты по экспериментальной эмбриологии уже довольно давно поняли, что у некоторых организмов процессы развития способны обеспечить образование хорошо сформированных адаптивных фенотипов даже при наличии значительных нарушающих воздействий извне — процесс, известный под названием гомеостаза развития. Так, например, еще в XIX в. Ганс Дриш (Hans Driesch) разрезал зародышей морского ежа на стадиях гаструлы и прегаструлы пополам (рис. 4.5) и получал из таких половинок вполне пропорциональных взрослых особей, хотя и несколько меньших размеров, чем обычные.
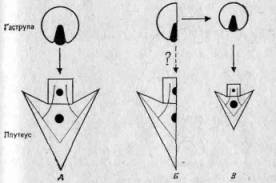
Рис. 4.5. Эксперименты Дриша на зародышах морского ежа. А. Нормальное развитие. Б. Гаст-рула, которую Дриш разрезал пополам, ожидая, что из нее разовьется «урод». В. Вместо ожидаемого урода из этой гаструлы в результате ее реорганизации развился маленький, но полный плутеус, способный в дальнейшем образовать нормальную взрослую особь.
2. Мутации в большинстве случаев рецессивны. В 20-х годах нашего века Морган (Morgan) заметил, что в природе все особи, принадлежащие к данному виду Drosophila, удивительно похожи друг на друга, и объяснял это тем, что все они обладают одним и тем же генотипом дикого типа; в сущности, это и предсказывал Дарвин (см. разд. 3.5). Добржанский (4), однако, показал, что подобное объяснение неверно; одинаковые на вид родительские особи могут иметь различные генотипы, поскольку генетическое разнообразие частично маскируется доминант-ностью. И в самом деле, само доминирование могло возникнуть именно по этой причине, то есть как проявление механизма, демпфирующего воздействие мутаций (см. разд. 3.7).
3. Эдмондс и Соуин (5) и Соуин и Эдмондс (10) изучали изменчивость строения дуги аорты у одомашненного кролика. На материале 3000 вскрытий они выделили всего лишь 20 типов строения, которые можно свести к 6 основным категориям (рис. 4.6). Эти 6 категорий встречаются с разной частотой. Такого рода наблюдения позволяют предполагать, что сложным системам органов свойственна ограниченная и не случайная изменчивость. Эдмондс и Соуин установили, что разные типы строения дуги аорты — результат различных скоростей роста передне-грудной области и осевого скелета.
4. При направленном отборе по отдельным признакам, проводимом в лаборатории, часто наблюдается следующая картина: после первоначальной быстрой реакции достигается определенный порог, и, несмотря на сильное давление отбора, дальнейшее изменение вызвать не удается (рис. 4.7). Подобная устойчивость фенотипа к сильному внешнему давлению отбора, возможно, обусловлена быстрым истощением изменчивости в популяции, подвергаемой отбору, и(или) участием внутренней, генетической и морфогенетической регуляции. Полагают, что последняя имеет важное значение.
Рис. 4.6. Вариации в строении дуг аорты у кроликов, выявленные на материале 3000 вскрытий (10). Существует 6 основных типов строения, к которым относятся 99,7% всей изменчивости. Указана частота каждого типа.
Рис. 4.7. Обычный тип реакции на направленный отбор. Объяснение см. в тексте.
5. Хоботок дрозофилы иногда превращается в усик, антенна— в ногу, голова — в грудной сегмент, проторакальный сегмент— в мезоторакальный и так далее. Морфологические изменения, в результате которых одна структура превращается в другую, называют гомоэотическими трансформациями. У насекомых такие трансформации встречаются довольно часто; при этом они возникают не беспорядочно, а в соответствии с определенными правилами: а) дорсальные структуры редко превращаются в вентральные, и наоборот; б) трансформации предсказуемы: трансформируемый орган (аутотип) обычно превращается лишь в один или несколько других органов (аллотипы— органы, которым он уподобляется в результате трансформаций); в) у дрозофилы в большинстве случаев наблюдаются трансформации в мезоторакальные структуры. Эти трансформации вызываются мутациями или тератогенными (изменяющими процесс развития) факторами. Результаты действия последних имитируют генетические эффекты, и их описывают под названием фенокопий. Эти гомоэотические трансформации иллюстрируют неслучайность и направленность морфологических трансформаций. Они позволяют предполагать, что существует некоторое ограниченное число траекторий развития, доступных организму после возникновения генетических или средовых возмущений.
У нас очень мало сведений о механизмах, стоящих за этими различными аспектами регуляции развития. Уоддингтон (12) назвал ее канализацией развития (каналы = траектории; Уоддингтон назвал их креодами) и представлял в виде мяча, перемещающегося по рельефному ландшафту (рис. 4.8). Мячик изображает продвижение развития. Средовые и генетические возмущения стремятся отклонить мячик с его пути; допускаемая степень такого отклонения зависит от глубины каналов, то есть от степени канализации. Один генотип может допускать развитие по одному из нескольких онтогенетических путей, а степень реализации каждого данного пути зависит от их относительной канализации. Какая-либо пертурбация, возникшая в среде, может выбить мячик из одного канала и столкнуть его в соседний канал. Вместе с тем такая пертурбация может привести к отбору на «углубление» альтернативного канала.
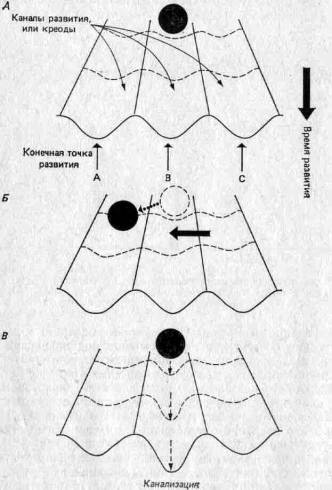
Рис. 4.8. Модель развития, предложенная Уоддингтоном. А. Мяч перемещается по каналу, или креоду, к некой конечной точке. Б. Возникающие в среде возмущения (черная горизонтальная стрелка) могут отклонить развитие, направив его к какой-то другой конечной точке. В. Отбор может углубить каналы и обеспечить преимущество одной конечной точки.
Это представляет собой одно из возможных объяснений наблюдения, что гомоэотические трансформации могут возникать под действием генетических изменений и (или) изменений в среде.
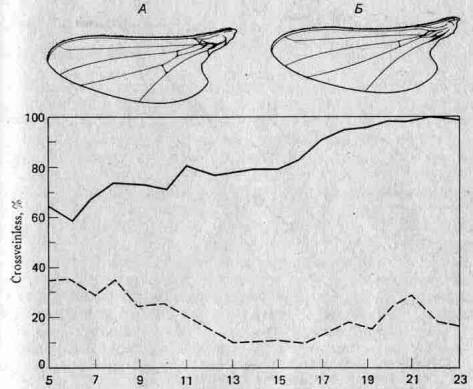
Рис. 4.9. Развитие у дрозофилы крыльев без поперечных жилок (crossveinless) под действием теплового шока. А. Нормальное крыло. Б. Крыло crossveinless; такое крыло может превратиться в нормальное в результате отбора среди мух с этим признаком после теплового шока (сплошная линия); штриховая линия — результаты отбора среди мух, которые не реагировали на тепловой шок.
Уоддингтон провел еще одно интересное исследование кана.-лизации. В обычных условиях у Drosophila melanogaster развивается крыло с нормальным набором поперечных жилок (рис. 4.9). Если подвергнуть мух во время развития тепловому шоку (40°С), то некоторые из этих жилок исчезают (то есть возникает состояние crossveinless; рис. 4.9). Более того, отбирая такие отклоняющиеся формы на протяжении нескольких поколений, Уоддингтон обнаружил, что крылья crossveinless могут возникать автоматически, без теплового шока. На первый взгляд создается впечатление, что мы здесь имеем дело с наследованием приобретенного признака; однако возможно и более правдоподобное неодарвинистское объяснение, которое иллюстрирует рис. 4.10. Генетический потенциал для признака crossveinless всегда имеется в виде «менее предпочтительной траектории».
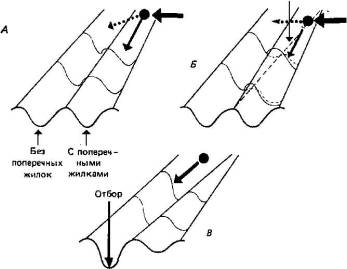
Рис. 4.10. Объяснение результатов, приведенных на рис. 4.9. А. Тепловой шок (черная горизонтальная стрелка) отклоняет мячик, направляя его на траекторию crossveinless. Б. Отбор понижает высоту «хребта» между траекториями, облегчая отклонение. В. Состояние crossveinless становится нормальным, а действие отбора углубляет (канализирует) эту траекторию.
Под действием теплового шока мячик перескакивает через холм на эту траекторию, и отбор (в данном случае — искусственный) приводит к уменьшению высоты холма и углублению альтернативного канала. Непосредственный эффект всего этого — создание фенокопий, а долговременный — реканализа-ция. Возможно, что такой же процесс участвовал в эволюции уплотненной кожи на подошвах ног у человека. Кожа обладает генетическим потенциалом к «уплотнению» при наличии соответствующего стимула, то есть стирания. Кожа на подошвах ног непрерывно стирается при ходьбе и поэтому уплотняется. Однако те индивидуумы, которые оказались способными наиболее эффективно реагировать на это (то есть те, у которых были «сравнительно низкие холмы» и более глубокие «каналы уплотненной кожи») могли иметь некое селективное преимущество. Таким образом, процесс уплотнения мог канализироваться, и теперь уплотненная кожа развивается без всякого стимула еще до того, как ребенок начинает ходить.
4.4. Заключения.
Тот факт, что мелкие генетические изменения могут ампли-фицироваться в процессе развития, приводя к более крупным фенотипическим эффектам, и что морфогенетические процессы часто проявляют устойчивость к нарушениям развития, создаваемым генетическими факторами и факторами среды, позволяет считать, что зависимость между генотипом и фенотипом носит нелинейный характер. Кроме того, изменчивость признаков, детерминируемых генами, ограничена и некоторые их изменения, по-видимому, более вероятны, чем другие. Следовательно, можно сказать, что возникновение генетически детерминированных фенотипических новшеств направляется фенотипом, и это ослабляет концепцию случайности в ее неодарвинистском понимании. Значение амплификации и канализации для эволюционной теории пока еще весьма неясно. Несомненно, однако, что как в адаптационистской, так и в генетической программе эволюционной биологии необходимо учитывать ограничения, налагаемые развитием.
4.5. Рекомендуемая литература.
Общим введением в проблемы «эволюционной биологии развития» могут служить книги Уоддингтона (12), Гоулда (6), Ол-берча и др. (2) и Олберча (1). Это сравнительно новая и сложная область, а поэтому читателю не следует рассчитывать, что он сможет найти в литературе достаточно последовательное и полное ее изложение.
Глава 5. МАКРОЭВОЛЮЦИЯ.
5.1. Введение.
Большая часть предыдущих глав была посвящена вопросу о том, каким образом организмы достигают соответствия со своей средой и постоянно поддерживают его (иногда это называют анагенезом). В этой последней главе мы займемся другим важным эволюционным процессом — происхождением разнообразия (иногда его называют кладогенезом). Макроэволюция— термин, означающий эволюционное развитие таксономических единиц видового и надвидового уровней. Эволюция этих единиц составляет основу создания таксономического разнообразия, и поэтому в рамках данной книги макроэволюция и кладо-генез будут использоваться как синонимы. Мы начнем с рассмотрения того, какие типы таксбнов можно различать и как их можно классифицировать.
5.2. Таксономические категории и система классификации.
Человек всегда стремился давать названия окружавшим его животным и растениям и обычно обращал внимание на существующую в живом мире иерархию таксонов. Последнее означает, что группы организмов, сходных по нескольким важным признакам, можно разделить на группы, обладающие этими и рядом других общих признаков, а эти группы можно подразделить в свою очередь на подгруппы и так далее. Линней рассматривал это как некую шкалу упорядоченности, приданную природе творцом, и представил ее в виде ряда категорий, расположенных в иерархическом порядке. Его система имела следующий вид:
ЦАРСТВО
КЛАСС
ОТРЯД РОД ВИД
Чем ниже место, занимаемое категорией в этом ряду, тем больше число общих признаков у входящих в нее подгрупп. С течением времени было создано много дополнительных категорий, но в основе системы классификации живых организмов все еще прочно лежит линнеевская иерархия. Большинство современных биологов различают следующие категории:
В ботанике и микробиологии типу соответствует отдел, а отряду—-по« рядок.
ЦАРСТВО
ТИП
КЛАСС
отряд
семейство .
РОД
вид
Все эти категории можно дробить дальше; так, например, на уровне отряда можно различать надотряды, отряды, подотряды и инфраотряды; подобные же категории можно создавать на любом другом уровне. В табл. 5.1 дана полная характеристика положения в системе медоносной пчелы и человека.
Таблица 5.1. Положение в системе медоносной пчелы и человека.
| Категория | Медоносная пчела | Человек |
| Тип | Arthropoda | Chordata |
| Подтип | Vertebrate | |
| Класс | Insecta | Mammalia |
| Подкласс | Pterygota | Eutheria |
| Отряд | Hymenoptera | Primates |
| Подотряд | Apocrita | Simiae |
| Род | Apis | Homo |
| Вид | mellifera | sapiens |
Примечание: название каждого вида слагается из двух слов — его родового и видового названий. Так,
Человек = Homo sapiens Медоносная пчела = Apis mellifera.
Такую систему обозначений называют биномиальной, и она также была введена Линнеем. Родовое название принято писать с прописной буквы, а видовое — со строчной; оба названия выделяют курсивом.
Для Линнея иерархическая система живых организмов отражала упорядоченность, которую заложил творец в свое создание. Для современных биологов это упорядоченность, в основе которой лежат общность происхождения и эволюционное родство. Основной единицей в том и другом случае служит вид: для Линнея — потому что он верил, что это единица, сотворенная богом, а для эволюционистов — потому что это та единица, на которую действует естественный отбор. В принципе вид можно теперь объективно определить как группу особей, способных обмениваться друг с другом генами, то есть скрещиваться между собой; тем самым эта группа обладает общим генофондом, доступным действию естественного отбора. Такое определение известно под названием биологической концепции вида. На практике однозначно идентифицировать биологические виды не удается, особенно если речь идет о мертвых и фоссилизирован-ных индивидуумах; поэтому обычно пользуются более прагматическим определением: вид — это категория, в которую систематики объединяют индивидуумов. Во всяком случае, современные биологи согласны в том, что эти группы эволюционировали от предковых видов, так что все современные формы связаны более или менее близким родством; поэтому они стараются распределить виды по группам, которые бы отражали эти эволюционные родственные связи.
Эволюция порождает явления двух разных типов, которые необходимо принимать во внимание при создании систем классификации: а) определенный порядок происхождения от предков; б) ту или иную степень дивергенции от предков. В общем, первое обусловливает черты сходства, а второе — различия. Представим себе три вида, возникающие одновременно от общего предка. У двух из них развились только мелкие различия, тогда как третий довольно заметно дивергировал от двух других (например, у него возник некий существенный новый признак). Если классифицировать эти формы на основании первого принципа, то следует считать, что все три вида связаны одинаковой степенью родства; если же применить второй принцип, то вид, обладающий новым признаком, следует отделить от двух других видов. Существует три разных взгляда на значимость этих критериев.
1. Кладизм (5, 7)
Этот подход основан на принципах, сформулированных энтомологом В. Хеннигом в 1950 г. (но см. (7)); главная роль в нем отводится генеалогии, то есть ветвящимся последовательностям, как единственному объективному способу установления систематического положения. (Слово «кладизм» происходит от греч. «кладос» — ветка или молодой побег.) Все члены данного так-сона (клады) должны брать начало от одного и того же предка, и организмы следует группировать только в соответствии с последовательностью их отделения от общего предка. Этот анализ обычно бывает основан на допущении, что все виды, существующие в любой данный момент времени, связаны родственными узами благодаря происхождению от общих предковых форм (рис. 5. 1, А, I но не //), не будучи сами предками каких-либо других существующих в это время видов. Следовательно, все современные виды можно объективно расположить в некой последовательности, строго основанной на генеалогическом родстве, и такое расположение можно схематически представить в виде кладограммы (рис. 5.1, А, III). Кладограммы строятся на основе наблюдаемого, или предполагаемого, числа признаков, встречающихся одновременно у всех групп. Однако к сходству как таковому следует относиться с осторожностью, так как может оказаться, что оно всего лишь отражает конвергенцию (развитие у различных структур внешнего сходства вследствие одинаковой функции; например, крылья летучих мышей, птиц и насекомых). Эволюционные родственные связи могут основываться только на признаках, сходство которых обусловлено тем, что они унаследованы от общего предка; такие признаки называют гомологичными. Чем больше общих гомологичных признаков имеется у двух видов, тем ближе родство между ними. Два вида, состоящие в самом тесном родстве, обладают гомологичными признаками, не встречающимися ни у каких других видов. Однако мы не смогли бы далеко уйти, если бы строили систему классификации на основе фундаментальных признаков, свойственных большинству организмов (таких, например, как многоклеточ-ность или даже пятипалое строение конечности у позвоночных), потому что, по определению, они действительно имеются у большинства организмов. Такие признаки называют общими примитивными признаками (иногда их называют симплезио-морфньши признаками). Кладисты утверждают, что ветвящиеся последовательности можно выделять лишь в том случае, если удается идентифицировать общие специализированные признаки (иногда их называют синапоморфными) — гомологичные сходства, наличествующие в самой точке ветвления. Возьмем, например, кошку, льва, тюленя и человека. У них много общих примитивных признаков (многоклеточность, кровь, пятипалые конечности и т. п.). Однако крупный головной мозг и бинокулярное зрение — это общие специализированные признаки, свойственные только приматам, и они определяют первую точку ветвления, отделяющую человека. У кошек и львов больше общих специализированных признаков (например, утрата верхних коренных, превращение верхних предкоренных в хищнические зубы), чем у каждого из них с тюленем, и это определяет нашу вторую точку ветвления. Кладограмма, получающаяся в результате такого анализа, изображена на рис. 5.1.А, III, где, а — человек, b — тюлень, с — лев, а — кошка.
И один последний вопрос: как кладист отличает примитивные признаки от специализированных? Один из важных методов состоит в так называемом сравнении за пределами группы (out-group comparison). Если признаки сходства между видами в пределах рассматриваемой группы обнаруживаются также у видов, не принадлежащих к ней, то это скорее примитивные, чем специализированные признаки. Потенциально возможно провести сравнение обширнейшего ряда видов, а именно это мы подсознательно и проделываем, когда называем такие признаки, как многоклеточность, примитивными. Однако обычно имеет практический смысл в известной мере ограничить подобного рода исследования, и интуитивно наиболее разумный подход состоит в том, чтобы сравнивать виды, входящие в данную группу, с другими сходными видами, не относящимися к ней.
2. Фенетика (16, 17)
Биологи, принадлежащие к этой школе, считают, что объективно классифицировать организмы в соответствии с их генеалогией невозможно главным образом из-за невозможности твердо отличать гомологичные признаки от конвергентных. Вместо этого они концентрируют внимание на всех чертах сходства и пытаются на их основе (то есть на основе степени общего сходства) создать объективную систему классификации организмов. Они настаивают на том, что если учитывать достаточное число признаков, то гомологичные признаки будут преобладать над конвергентными. Однако гарантировать это a priori нельзя. Другие приверженцы фенетики утверждают, что они стремятся лишь к тому, чтобы создать удобные системы классификации, подобные тем, которыми пользуются в библиотеках для классификации книг, и что эти системы не обязательно должны отражать эволюционные взаимосвязи. Для проведения фенетического анализа (то есть оценки степени общего сходства) часто применяют хитроумные математические методы, выражая полученные результаты в виде фенограмм (рис. 5. 1Д).
3. Эволюционная систематика (13)
Рис. 5.1. А. Кладисты обычно исходят из допущения, что рассматриваемые виды связаны между собой так, как это показано на схеме /, но не так, как на схеме //. На схеме III показана кладограмма, основанная на последовательных ветвлениях. Вопросительный знак у оси времени стоит потому, что кладограммы можно интерпретировать по крайней мере двумя способами. Согласно одной интерпретации, они представляют собой эволюционные древеса, на которых предки точно не указаны, а поэтому вводится время (классическая кладистика); согласно другой — они изображают типы родственных взаимоотношений, а точки ветвления соответствуют синапо-морфным признакам, так что время не вводится (трансформированная кладистика). Классическую кладистику интересуют эволюционные процессы, а трансформированную — таксономические картины. Конечно, те и другие должны быть как-то связаны между собой, однако некоторые кладисты предпочитают не высказываться относительно точного характера этой связи. Б. Ступенчатое построение фенограммы. Буквы обозначают виды. Числа в матрицах — коэффициенты общего сходства (в диапазоне от 0 — отсутствие сходства до 1 — полное сходство), выведенные на основании всех измеренных признаков. На ступени / матрица содержит коэффициенты сходства для пяти видов. Построение фенограммы начинают с поиска основных пар (для каждого члена такой пары коэффициенты сходства максимальны). Эти основные пары соединяют на соответствующих уровнях оси, по которой отложены коэффициенты сходства. На ступени // каждая основная пара рассматривается как одна единица, и коэффициенты сходства здесь представляют собой средние по всем отдельным видам, то есть 0,575= (0,6+0,4+ + 0,7+0,6)/4. Здесь снова следует найти основные пары и произвести соответствующие соединения на схеме. На ступени /// продолжают и завершают эту процедуру (по Luria S. Е. et al., A. View of Life, Benjamin, 1981). В. Филогенетическое древо, в котором сделана попытка учесть как последовательность ветвления (а тем самым сроки), так и степень дивергенции. Время обычно пытаются отложить как можно точнее на вертикальной оси; горизонтальная ось отражает дивергенцию обычно на основании субъективных оценок, а поэтому довольно неточно.
В эволюционной систематике классификация основана на сочетаний генеалогии и степени общего сходства и различия, отражающего уровень дивергенции. Никаких объективных правил для этого не существует, и принятие того или иного решения в значительной мере зависит от опыта систематиков в оценке относительной значимости различных критериев. Полученные результаты представляют в виде филогенетических древес (рис. 5.1.В).
Некоторые из различий, существующих между этими школами, можно проиллюстрировать на следующих примерах, относящихся к птицам и рептилиям. Как птицы, так и крокодилы происходят от общей предковой группы архозавров, в которую входили динозавры. Какая-то еще более древняя рептилия была общим предком черепах, ящериц и змей, крокодилов и птиц. Поэтому у птиц и крокодилов больше общих признаков, чем у крокодилов и других ныне существующих рептилий.
Однако после того, как птицы отделились от общего ствола, они развивались быстрее, чем крокодилы, и теперь они обладают рядом чрезвычайно своеобразных признаков — способностью к полету, перьевым покровом, гомойотермностью и т. п. Поскольку птицы сильнее дивергировали от предкового ствола, чеш крокодилы, эволюционные систематики отделяют их от всех других рептилий и помещают в отдельный класс — Aves, a всех остальных ныне существующих рептилий, в том числе крокодилов, объединяют в класс Reptilia (рис. 5.1,Б). Фенетики строят аналогичную систему на основе морфологического сходства. Что же касается кладистов, то они при построении систем классификации строго придерживаются генеалогического родства, и поэтому в их кладограммах птицы связаны с крокодилами более тесным родством, чем крокодилы — с черепахами, змеями и ящерицами.
Итак, подводя итоги, можно сказать, что какого-то оптимального способа построения биологической системы классификации, которая отражала бы эволюционные связи, по-видимому, не существует. Кладисты строят ее объективно на основе одной лишь генеалогии, но пренебрегают важными сведениями о дивергенции. Фенетики строят ее объективно, основываясь на сходствах, но неизбежно путают при этом гомологичные и конвергентные признаки. Представители эволюционной систематики пытаются учитывать одновременно и генеалогию, и дивергенцию, но не могут делать это вполне объективно.
Однако независимо от того, какой мы придерживаемся методологии, производя отбор групп однородных таксонов, следует найти их место в линнеевской иерархии, то есть возвести их в определенный ранг, как, например, отряд или семейство. Кладисты часто вводят новый ранг после каждой точки ветвления в кла-дограмме. Эволюционные систематики, однако, судят о ранге таксонов по степени их дивергенции от общего предка, нередко приписывая разные ранги сестринским группам. Определение ранга — процесс крайне субъективный, а поэтому он вызывает многочисленные разногласия и подвержен всевозможным изменениям и уточнениям.
5.3. Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции.
Так называемая синтетическая теория эволюции пытается объяснить происхождение таксономического разнообразия, то есть макроэволюционные явления, с точки зрения принципов неодарвинизма, то есть микроэволюционных процессов. Важными вехами такого подхода служат книги Добржанского (4), Хаксли (8), Майра (12) и Симпсона (15).
Синтетическая теория строит свое объяснение видообразования, на основных принципах неодарвинизма, а именно:
1) источником изменчивости служат точковые мутации, в особенности мутации структурных генов;
2) эволюционное изменение представляет собой результат изменения частоты генов;
3) направление этих изменений частоты определяется естественным отбором.
Таким образом, традиционно считается, что популяции некоего вида становятся физически изолированными, после чего обмен генами между ними прекращается. В результате этой изоляции возникают группы, представляющие собой более или менее случайные выборки из первоначального генофонда. Возможно также возникновение неслучайных выборок, как, например, в случае эффекта основателя, однако, согласно приверженцам синтетической теории, это бывает редко. Изоляты становятся адаптированными к тем условиям, в которых они оказались, так что их генные частоты начинают различаться, и в конце концов популяции оказываются настолько разными, что даже в случае разрушения изолирующих преград скрещивание между ними становится невозможным — иными словами, они превращаются в настоящие виды (рис. 5.2). Решающим моментом для подобного процесса видообразования служит возникновение изоляции, за которым следуют медленные и постепенные изменения. Это схематически показано на рис. 5.2.
Интерпретация видообразования, как процесса постепенного, сталкивается с двумя проблемами: одна из них носит фактический характер, а другая —логический. Что касается фактов, то в палеонтологической летописи часто нельзя обнаружить постепенных изменений. Многие виды, казалось бы, остаются неизменными на протяжении миллионов лет, а затем внезапно исчезают, сменяясь какими-то другими формами, существенно отличающимися от прежних, но явно родственными им. Хорошим примером служит группа пресноводных моллюсков из поздне-кайнозойских отложений озера Туркана, на севере Кении, описанная Уильямсом (21). Как показали проведенные измерения, за последние 5 млн. лет популяции 16 видов изменялись весьма незначительно. Однако у 5 видов, мало изменявшихся в течение почти всего этого времени, обнаружены очень быстрые изменения за периоды, равные примерно 50 000 лет. Со стороны логики можно привести то, что Майварт (Mivart) назвал дилеммой стадии зарождения полезных структур. Если эволюция структур происходит постепенно, то какое значение имеют структуры с промежуточными признаками? Какую пользу, например, может принести рептилии частично сформированное крыло (см. также разд. 1.5.)?

Рис. 5.2. Видообразование как постепенный процесс. После возникновения изоляции (горизонтальная стрелка) изолят (вид В) постепенно дивергирует от «родительского» вида А.
Сторонники синтетической теории (или сторонники постепенной эволюции) пытаются справиться с фактической стороной проблемы, утверждая, что отсутствие постепенных изменений — явление не столь частое, как это иногда полагают, и что его можно объяснить неполнотой палеонтологической летописи. Накопление осадков и захоронение костей — процесс капризный, особенно если речь идет о группах с низкой плотностью популяций, а поэтому промежуточные формы должны быть редкими. Для объяснения дилеммы Майварта они привлекли преадапта-цию: промежуточные структуры выполняют определенные роли, для которых они хорошо подходят, но чисто случайно они соответствуют и другим ролям, которые могут играть после дальнейшего усовершенствования. Складка кожи между передней конечностью и туловищем могла развиваться у рептилий в качестве приспособления для терморегуляции, а затем найти применение при ускальзывании от врага и в конечном итоге дать начало крылу. Подобным же образом воздушный мешок у рыб возник не для того, чтобы они когда-нибудь могли заселить сушу, а в качестве дополнительного органа дыхания, используемого в водной среде. Однако, после того как воздушные мешки образовались, они стали использоваться в качестве легких в связи с такими изменениями в условиях среды, как пересыхание водоемов.
5.3.1. К вопросу об изоляции и видообразовании
Выше подчеркивалось, что изоляция — это важная предпосылка для видообразования. В настоящее время известно, что существует несколько возможных способов возникновения изоляции, а следовательно, и способов видообразования. Обзор их дал Буш (1).
Поток генов (обмен генами) между популяциями может быть подавлен какой-либо физической или географической преградой. Этот процесс называют аллопатрическим видообразованием. В принципе возможен также альтернативный процесс — подавление обмена генами в популяциях, обитающих в одной и той же области — например если носители благоприятных мутаций спариваются преимущественно между собой и если отбор в пользу этих мутаций достаточно интенсивен, чтобы возобладать над обменом генами, создаваемым спариваниями с более часто встречающимися родительскими формами. Этот процесс носит название симпатрического видообразования. И наконец, даже при наличии некоторого обмена генами между частично изолированными популяциями в областях, в которых они примыкают одна к другой, возможна, опять-таки в принципе, дивергенция признаков под действием отбора, если обмен генами относительно слабый, а различия в давлениях отбора относительно велики. Этот процесс известен под названием парапатрического видообразования и может происходить относительно часто у организмов с низкой подвижностью ( = слабой способностью к расселению), таких, как растения и наземные моллюски.
Если значение аллопатрического видообразования для эволюции у большинства биологов не вызывает сомнений, то относительно эволюционной роли симпатрического и парапатрического видообразования идет много споров. При этом вновь подчеркивается важность демовой структуры (как, например, на рис. 2.9,Д) для процесса видообразования.
Следует также привлечь внимание к различию между географической изоляцией (то, что было описано под названием аллопатрии) и изолирующими механизмами. Изолирующие механизмы— это различия в признаках между популяциями, препятствующие скрещиваниям между ними и сохраняющие целостность вида, когда популяции встречаются вместе. Эти механизмы многочисленны и разнообразны, и среди них есть как преко-пуляционные изолирующие механизмы (например, морфологические и поведенческие признаки, препятствующие копуляции самцов и самок), так и посткопуляционные изолирующие механизмы (например, несовместимость гамет, которая препятствует оплодотворению яиц или приводит к стерильности потомства, как в случае мулов).
5.4. Альтернативная точка зрения — прерывистая эволюция.
Существует и другая точка зрения, альтернативная изложенной в предыдущем параграфе теории постепенной эволюции. Палеонтологическая летопись, возможно, не столь неполна, как это предполагается. Быть может, эволюция на самом деле слагается из кратковременных периодов быстрого изменения, за которыми следуют длительные периоды равновесия. Эта теория прерывистого равновесия обладает тем преимуществом, что позволяет объяснить то, что мы, по-видимому, наблюдаем, и разрешить дилемму Майварта, поскольку всякая необходимость в существовании промежуточных форм при этом просто отпадает. Она подразумевает возможность быстрого возникновения крупных изменений формы и функции. Дарвин и неодарвинисты полагали, что подобная точка зрения подорвет доверие к их теориям, поскольку крупные изменения в преобладающем большинстве случаев должны быть гибельными, то есть несовместимыми с фенотипом (см. разд. 1.5).
Рис. 5.3. Видообразование как прерывистый процесс. Вслед за быстрым видообразованием (горизонтальная стрелка) наступает период, во время которого изменений происходит мало.
В сущности, это соответствует также теории прерывистой эволюции, так как ее сторонники считают, что длительные периоды застоя отражают неспособность большей части мутационных изменений войти в состав фенотипа, то есть ограничения, связанные с коадаптацией и интеграцией, противодействуют изменению. Время от времени, однако, одна из таких изменений, которое Гольдшмидт в своей книге «Материальные основы эволюции» (Goldschmidt, The Material Basis of Evolution, Yale University Press, 1940) назвал «подающими надежды чудовищами», может оказаться совместимым с существующим адаптивным комплексом. Теперь мы знаем, что возникновение и интеграция таких крупных мутационных изменений согласуется с процессами развития (гл. 4). Возможен и альтернативный путь: возникновение резкого сдвига в сторону от генофонда родительской популяции в результате образования неслучайной выборки на ранних стадиях изоляции. Этот путь считается наиболее вероятным для мелких периферических изоля-тов больших популяций. Создающаяся при этом картина прерывистой эволюции представлена на рис. 5.3. (Обратите внимание на сходство между этим рисунком на рис. 5.1 А/7/, то есть кла-дограммой. Кладисты часто относятся весьма благосклонно к теории прерывистого равновесия, а сторонники последней нередко поддерживают кладизм. Это связано с тем, что при кладисти-ческом анализе принимается допущение о периодах быстрого видообразования с незначительной дивергенцией между точками ветвления.)
Рис. 5.4. Направление эволюции, создавшееся в результате дифференциального видообразования и выживания видов, а не в результате постепенных изменений. Виды возникают внезапно, а затем почти не изменяются. Сдвиг вправо обусловлен тем, что в этом направлении видообразование происходит чаще, а также тем, что виды с такими морфологическими признаками выживают дольше. По-видимому, возможно преобладание любого из этих процессов, то есть как дифференциального видообразования, так и вымирания.
Таким образом, как и традиционный неодарвинизм, теория прерывистого равновесия представляет себе начальный источник изменчивости как случайный по отношению к направлению-эволюции, но рассматривает эту изменчивость как сопряженную с изменениями гораздо большего масштаба. Кроме того, вследствие этого и последующего застоя теория прерывистого равновесия рассматривает источник изменчивости, то есть изоляцию или периоды видообразования, как самые важные элементы, определяющие направление и скорость эволюции (возврат к взглядам ранних менделистов?).
Но если это так, то в результате каких же процессов возникают адаптации и как создаются эволюционные направления, если и то, и другое — черты, ярко выраженные как в палеонтологической летописи, так и у ныне существующих видов? Для сторонников прерывистой эволюции ответом будет отбор видов (термин, созданный Стенли (18)). Согласно этой точке зрения, эволюционные направления могут возникнуть: 1) потому что некоторые случайно образовавшиеся изоляты более устойчивы, чем другие, либо (а) потому, что они лучше приспособлены в обычном смысле, либо (б) вследствие экологических условий той местности, в которой они оказались изолированными (например, случайное отсутствие какого-либо прожорливого хищника); и (или) 2) потому что у некоторых видов в результате их организации или экологии видообразование происходит чаще, чем у других. Эти возможности проиллюстрированы на рис. 5.4. Как в случае 1, так и в случае 2 создаются эволюционные направления, но возникающие при этом адаптации, особенно в случаях 16 и 2, вряд ли окажутся такими совершенными, как те, которые могут возникнуть в результате более обычного постепенного видообразования.
В ее современной форме теорию прерывистого равновесия отстаивают, в частности, Гоулд и Элдридж (6) и Стенли (18, 19). Однако, как уже отмечалось, она уходит своими корнями в работы Гольдшмидта (см. выше), а также де Фриза (3). Т. X. Хаксли, один из самых верных союзников Дарвина, был также склонен признавать значительные скачки в процессе эволюции — к вящему неудовольствию самого Дарвина!
5.5. Сравнение теорий постепенной и прерывистой эволюции.
Для того чтобы показать более конкретно различие между постепенной и прерывистой эволюцией в плане эволюционных направлений и адаптации, рассмотрим классический пример эволюции современной лошади, далеким предком которой был Hyracotherium — трехпалое существо величиной не больше собаки. Палеонтологическая летопись свидетельствует, по-видимому, о постепенном изменении размеров тела и формы, приведшем в конечном счете к хорошо знакомому нам Equus. Классическое объяснение с позиций постепенной эволюции сводится к прогрессивному изменению под действием естественного отбора (см. разд. 3.2). В противоположность этому теория прерывистого равновесия объясняет направления в эволюции лошадей как результат асимметрии генеалогического древа. Эволюционная история лошади рассматривается как видообразовательная линия, в которой возникали некоторые новые виды, отличавшиеся большими размерами тела и сокращением числа пальцев, а также другие виды с совершенно иными морфологическими изменениями. Виды с более крупным телом и редуцированными пальцами оказались более удачливыми, чем виды с более примитивными признаками — большим числом пальцев и меньшими общими размерами, — и это привело к асимметрии генеалогического древа. С течением времени «центр тяжести» явно склоняется в сторону более крупного вида с одним пальцем (как, например, на рис. 5.4).
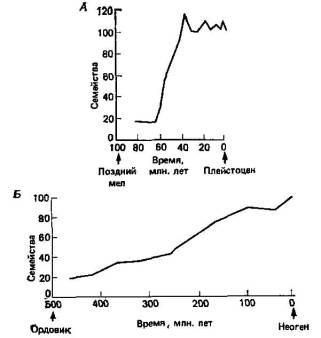
Рис. 5.5. Разнообразие семейств млекопитающих (А) и двустворчатых моллюсков (Б) на протяжении длительных периодов времени (19).
В табл. 5.2 перечислены главные различия между теориями постепенной и прерывистой эволюции. Таблица 5.2. Сравнение гипотез постепенного и прерывистого видообразования
| Фактор | Постепенное видообразование | Прерывистое видообразование |
| Изоляция | Случайный процесс, иногда неслучайный обычно аллопатрический | Изолированная подгруппа часто имеет неслучайный генетический состав; может быть симпатрическим или парапат-рическим |
| Источник изменчивости | Главным образом точковые мутации | Главным образом макромутации (то есть мутации в первоначальном смысле) |
| Отбор | Внутри популяций | Между популяциями (или видами) |
| Дивергенция | Медленная и непрерывная (и пропорциональная числу поколений) | Быстрая (непропорциональная числу поколений), но при отсутствии изменений в течение длительных периодов |
| Ограничения | Не очень существенные | Очень существенные |
| Адаптации | Очевидные и совершенные | Не столь очевидные и менее совершенные |
5.6. Проверка двух теорий эволюции.
Стенли (18,19) рассмотрел разного рода данные, которые можно использовать для сравнительной оценки теорий постепенной и прерывистой эволюции. К наиболее общим соображениям относится ожидание, что скорость макроэволюции (то есть скорость возникновения семейств и отрядов, отражающих крупные морфологические изменения) должна быть: 1) согласно модели постепенной эволюции, пропорциональна времени или, точнее, числу поколений, на протяжении которых могло развиваться разнообразие; и 2) согласно модели прерывистой эволюции, пропорциональна степени расщепления, то есть видообразования, которое имело место. Если удастся оценить как время, так и скорость расщепления, то тогда мы в принципе сможем провести различие между этими двумя гипотезами. На этой основе Стенли проводит сравнение между млекопитающими и двустворчатыми моллюсками. У млекопитающих скорость видообразования высокая и радиация в пределах группы, в результате которой возникло примерно 100 семейств, происходила в течение менее чем 30 млн. лет. У двустворчатых моллюсков, напротив, видообразование протекало медленно и соответственно происходила медленная радиация на протяжении более чем 300 млн. лет (рис. 5.5). Более того, среди двустворчатых моллюсков у одной группы сердцевидок (сем. Cardiidae) так называемого понтического ряда, изолировавшегося из Средиземноморья в Каспийском море, видообразование происходило быстрее, чем у ее близких родичей, и за менее чем 3 млн. лет эта группа достигла такого морфологического разнообразия, что ее теперь делят на четыре подсемейства.
Следствие, вытекающее из всех этих рассуждений, состоит в том, что в группах, в которых на протяжении длительных периодов геологического времени происходило мало эволюционных изменений, видообразование, очевидно, было незначительным. И действительно, так, по-видимому, обстояло дело со многими так называемыми живыми ископаемыми. Хорошим примером служат двоякодышащие рыбы. Быстрые морфологические изменения происходили у них только в период быстрого видообразования, в середине палеозоя, вскоре после их появления. После этого скорость видообразования резко понизилась, и вместе с ней упала и скорость морфологических изменений. Другим примером служат аллигаторы, строение которых в процессе эволюции изменялось мало, которые представлены в настоящее время всего двумя видами и никогда не отличались большим числом видов. Наконец, известен лишь один ныне живущий вид трубкозубов, а в палеонтологической летописи, вплоть до начала миоцена, к которому относятся самые ранние находки представителей этой группы, обнаружено лишь очень небольшое число их линий.
Все это свидетельствует в пользу теории прерывистой эволюции. Однако имеющиеся данные нельзя считать решающими, так как их анализ зависит главным образом от разделения ископаемых таксонов, которое может отражать в большей мере пристрастия систематиков, чем истинное течение эволюции; так, например, вопрос о том, какие группы видов образуют семейства, довольно субъективен. Даже соглашаясь принимать фактические данные, классический неодарвинист может заявить, что ряд факторов и, в частности, вариации в интенсивности естественного отбора перекрывают ожидаемую корреляцию между скоростью микроэволюции и временем, предсказываемую в пункте 1 в начале этого параграфа. Иными словами, неодарвинизм не настаивает на непременной постепенности. Он допускает возможность периодов быстрых изменений под действием интенсивного отбора и периодов медленных изменений под действием слабого отбора. Кроме того, застой в морфологии твердых частей тела — а именно они сохраняются в палеонтологической летописи—не обязательно означает застой в строении мягких частей, а также метаболических и поведенческих процессов. Таким образом, хотя фактические данные, очевидно, свидетельствуют в пользу теории прерывистой эволюции, они все еще далеко не окончательны, и вопрос остается открытым, вызывая горячие дебаты.
5.7. Была ли эволюция человека постепенной или прерывистой?
Креационисты (см. гл. 1) считали, что человек отделился от человекообразных обезьян с того момента, когда бог вдохнул в него душу! В ответ на это дарвинисты и неодарвинисты упорно настаивают на том, что человек произошел от человекообразных обезьян в результате постепенной прогрессивной эволюции. Исходя из этого, обычно принято помещать вид Homo sapiens в конец непрерывного ряда, после по крайней мере двух других видов (как, например, на рис. 5.6). Эти разные виды характеризуются следующим образом.
1. Ramapithecus (от Рама —индийское божество, и греч. пи-текос —обезьяна). Эта группа существовала примерно 8— 14 млн. лет назад. Она известна только по фрагментам нижней челюсти, которые позволяют считать, что лицевой отдел черепа у рамапитека был плоским, со слабо выступающей нижней
частью.
2. Австралопитек (от лат. austral —южный). Эта группа существовала 1,5—5 млн. лет назад. Лицевой отдел черепа у австралопитеков был плоским. Расположение черепа относительно позвоночника и костей задних конечностей относительно таза позволяет считать, что положение тела у него было вертикальным. Известны две главные группы австралопитеков: Australopithecus robustus с массивным черепом и крупными зубами, что свидетельствует о растительной диете, и группа, к которой относятся A. afarensis и A. africanus, с более изящным скелетом.
3. Homo habilis (человек умелый). Эта группа существовала 1,5—2 млн. лет назад. Ее представители были прямоходящими, имели относительно большой головной мозг и пользовались различными орудиями.
4. Homo erectus (человек прямоходящий). Эта группа существовала 1 млн. лет назад. У ее представителей походка была еще более сходна с нашей, мозг был крупнее, чем у Я. habilis, и они пользовались огнем.
5. Неандертальцы (от названия долины Неандерталь, где они были впервые найдены). Неандертальцы существовали примерно 0,5 млн. лет назад; они настолько сходны с современными людьми как по морфологии, так и по наличию определенной культуры с различными обрядами, что их считают одним из подвидов Homo sapiens.
Рис. 5.6. Две разные филогении, предложенные для основных таксонов гоминид (2). R — Ramapi-thecus; Aafa—Australopithecus afarensis; Aa — A. africanus; Ab—A. boisei; Ar — A. robustus; Hh — Homo habilis; He — H. erectus; Hs — H. sapiens.
Недавно были обнаружены два важных факта, которые возродили точку зрения о том, что эволюция человека — процесс прерывистый (19). Во-первых, было высказано мнение, что одновременно существовал не один, а несколько видов гоминид, а это указывает на то, что филогения рода Homo была кустистой, а не линейной. Во-вторых, полагают, что имеются данные, свидетельствующие о наличии относительно длительных периодов застоя в филогении как группы автралопитеков, так и гоминид (см., например, Australopithecus africanus, Homo habilis и Я. erectus на рис. 5.7), а это довольно удивительно, принимая во внимание высокую абсолютную скорость, характерную для эволюции человека.
Вместе с тем данные о перекрывании во времени между видами рода Homo не бесспорны, и если даже они верны, то не отвергают интерпретацию эволюции человека как процесса постепенного.
Рис. 5.7. Стратиграфические диапазоны гоминид (19). Прерывистыми линиями обозначены ископаемые остатки, предположительно отнесенные к тому или иному виду. Обозначения те же, что на рис. 5.6. N — неандерталец.
Ветвление могло просто увеличить число направлений, по которым эволюция человека протекала постепенно. Кроме того, что гораздо важнее, хотя разные таксоны гоминид резко различаются по некоторым морфологическим признакам, например по признакам черепа, количественные признаки, например размеры тела и емкость черепной коробки, изменяются почти непрерывно (рис. 5.8). Кроме того, по мнению некоторых антропологов, застой в пределах отдельных таксонов скорее кажущийся, чем реальный (2). Например, у самых ранних экземпляров Н. erectus лицевой отдел черепа более вытянутый и плоский, черепная коробка ниже, тоньше, менее прочна и емкость ее меньше, чем у более поздних Н. erectus.
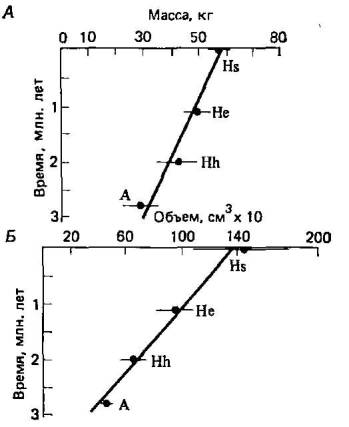
Рис. 5.8. Изменения среднего веса тела (Л) и объема черепной коробки (Б) у гоминид с течением времени (2). Обозначения те же, что на рис. 5.6.
5.8. Заключения.
Итак, остается все еще обширное поле для дискуссий между сторонниками прерывистой эволюции и традиционными неодарвинистами. Главная проблема, стоящая перед последними,— это,, вероятно, не скорость, с которой эволюционные изменения могут происходить в периоды быстрой эволюции, ибо при наличии соответствующих интенсивностей отбора быстрые изменения вполне осуществимы, а тот факт, что признаки могут оставаться неизменными на протяжении длительных периодов геологического времени. Так, например, Ланде (9) вычислил, что число элиминируемых отбором особей, необходимое для того, чтобы можно было объяснить изменения в некоторых скелетных структурах у кайнозойских млекопитающих, равно примерно 1 млн. Такое давление отбора настолько незначительно, что вполне могло возникнуть в результате совершенно случайных процессов. И тем не менее массовый дрейф генов, происходящий на протяжении миллионов лет, совершенно не входит в рамки неодарвинизма. Неодарвинисты могли бы также утверждать, что за кажущимся застоем скрывается интенсивный стабилизирующий отбор (см. разд. 2.3.3), но в таком случае нам придется допустить долговременный застой в условиях среды, что представляется в равной мере маловероятным.
В свою очередь сторонникам теории прерывистой эволюции надлежит прежде всего решить проблему случайности в скачкообразных изменениях. Прерывистые изменения должны возникать с одинаковой вероятностью во всех направлениях, включая и направление, в котором происходит эволюция. Это могло бы осуществляться путем дрейфа и эффекта основателя, однако широкое распространение этих процессов еще предстоит показать. Другая возможность состоит в том, что если основу прерывистой эволюции составляют крупные мутации, то они должны сообщать своим носителям способность к быстрому распространению, а это отличается от классического неодарвинизма только масштабом изменения.
Удастся ли найти ясное решение всех этих вопросов, опираясь на фактические данные, остается сомнительным. Кроме того, возможно, что эволюция одних групп протекала в соответствии с теорией прерывистого равновесия, а других —в соответствии с неодарвинистской моделью. Иными словами, вряд ли какая-либо одна модель эволюции окажется применимой ко всем случаям.
5.9. Рекомендуемая литература.
Хорошее изложение синтетической теории эволюции дают Майр и Провин (14). Для того чтобы оценить, как разные представители эволюционной биологии интерпретируют теорию прерывистого равновесия, читателю следует сравнить и противопоставить отчеты Левина (10) и Мэйнарда Смита (11) об одной и той же конференции по макроэволюции (Чикаго, октябрь 1980 г.). Один из номеров журнала New Scientist (15 апреля 1982, 194, № 1301), посвященный столетию со дня смерти Чарлза Дарвина, содержит статью Гоулда (Gould), отстаивающего прерывистую теорию, и статью Чарлзуорта (Charlesworth), направленную против этой теории, а также хорошую статью Стринджера (Stringer) об эволюции человека. Популярный очерк модели прерывистого равновесия дает Стенли (20).
ЛИТЕРАТУРА.
Глава 1
1. George W. Darwin, Fontana, London, 1982.
2. Gillespie N. С. Charles Darwin and the Problem of Creation, University of Chicago Press, Chicago and London, 1979.
3. Howard J. Darwin, Oxford University Press, Oxford, 1982.
4. Moore J. R. The Post-Darwinian Controversies, Cambridge University Press,. Cambridge, 1979.
5. Moorehead A. Darwin and the «Beagle», Hamilton, London, 1969.
6. Ridley M. Coadaptation and the inadequacy of natural selection, British J, of History of Science, 15, 45—68 (1982).
7. Ruse M. The Darwinian Revolution, University of Chicago Press, Chicago» and London, 1979.
8. Ruse M. Darwinism Defended, Addison-Wesley Publ. Co., Massachusetts, 1982.
9. Smith C. U. M. The Problem of Life, Macmillan Press Ltd., London, 1976.
Глава 2
1. Ayala F. J., Kiger J. A. Modern Genetics, Benjamin/Cummings Publ. Co., Menlo Park, California, 1980.
2. Berry R. J. Neo-Darwinism, Edward Arnold, London, 1982.
3. Charlesworth B. Evolution in Age-structured Populations, Cambridge University Press, Cambridge, 1980.
4. Cook L. M. Coefficients of Natural Selection, Hutchinson University Library, London, 1971.
5. Crow J. F. Genes that violate Mendel's rules, Scientific American, 240, 104—113 (1979).
6. Falconer D. S. Introduction to Quantitative Genetics, 2nd edn., Longman, London and New York (1st edn., Oliver and Boyd, 1960), 1981.
7. Gale J. S. Population Genetics, Blackie, Glasgow and London, 1980.
8. Ludovici L. J. The Chain of Life, Phoenix House Ltd., London, 1963.
9. Ricklefs R. E. Ecology, 2nd edn., Nelson, Middlesex, 1980. (Имеется перевод первого издания: Риклефс Р., Основы общей экономии. — М.: Мир, 1979.)
10. Roughgarden J. Theory of Population Genetics and Evolutionary Ecology: An Introduction, Macmillan Publ. Co., N. Y., 1979.
11. Turner J. R. G. Changes in mean fitness under natural selection. In: Mathematical Topics in Population Genetics (K. Kojima, ed.), Springer, Berlin, pp. 33—78, 1970.
12. Watson J. D. Molecular Biology of the Gene, 3rd edn., W. A. Benjamin Inc., California, 1976. (Имеется перевод: Уотсон Д., Молекулярная биология гена. — М.: Мир, 1979.)
13. Watson J. D. The Double Helix: A Norton Critical Edition (G. S. Stent. ed.), W. W. Norton, New York, 1980. (Имеется перевод: Уотсон Д., Двойная спираль. — М.: Мир, 1969.)
14. Watson J. D., Tooze J. The UNA Story, W. H. Freeman and Co., San Francisco, 1981.
15. Woods R. A. Biochemical Genetics, Chapman and Hall, London and New York, 1980.
16. Zimmering S., Sandier L., Nicoletti B. Mechanisms of meiotic drive, Ann. Rev. Genetics, 4, 409—436.
Глава 3
1. Alexander R. McN. Optima for Animals, Edward Arnold, London, 1982.
2. Bell G. The Masterpiece of Nature, Groom Helm Publishers, London, 1982.
3. Blower G. Age-structure of millipede populations in relation to activity and dispersion. In: The Soil Ecosystem (J. G. Sheal, ed.), Systematics Association Publ. No. 8, pp. 209—216, 1969.
4. Cain A, L, Sheppard P. M. Natural selection in Cepaea, Genetics, 39, 89— 116 (1954).
5. Calow P., Townsend C. R. Energetics, ecology and evolution. In: Physiological Ecology. An Evolutionary Approach to Resource Use (C. R. Townsend and P. Calow, eds.), Blackwell Scientific Publications, Oxford, pp. 3—19, 1981.
6. Calow P., Woollhead A. S. The relation between ration, reproductive effort and age-specific mortality in the evolution of life-history strategies — some observations on freshwater triclads, J. Anim. Ecol., 46, 765—781 (1977).
7. Cohn D. L. Optimal systems. I. The vascular system, Bull. Math. Biophys.,
16. 59—74 (1954).
8. Cohn D. L. Optimal systems. II. The vascular system, Bull. Math. Biophys.,
17. 219—223.
9. Cole L. C. The population consequences of life history phenomena, Quart. Rev. Biol., 29, 103-137 (1954).
10. Dames P. S. Physiological ecology of Patella I. The effect of body size and temperature on metabolic rate, J. Marine Biol. Ass., U. K., 46, 647—658 (1966).
11. Davies P. S. Physiological ecology of Patella III. Desiccation effects, J. Marine Biol. Ass., U. K., 49, 291—304 (1969).
12. Dawkins R. The Selfish Gene, Oxford University Press, Oxford, 1976.
13. Dawkins R. The Extended Phenotype, W. H. Freeman and Co., Oxford and San Francisco, 1982.
14. Dobzhansky Th. Genetics and the Origin of Species, Columbia University Press, N. Y., 1937.
15. Fisher R. A. The Genetical Theory of Natural Selection, Clarendon Press, Oxford, 1930.
16. Ford E. B. Ecological Genetics, Chapman and Hall, London, 1975.
17. Gilpin M. E. Group Selection in Predator-Prey Communities, Princeton University Press, Princeton, N. J., 1975.
18. Gould S. J., Lewontin R. C. The spandrels of San Marco and the Panglos-sian paradigm: a critique of the adaptationist programme, Proc. Roy. Soc. Lond., 205B, 581—598 (1979).
19. Hamilton W. D. The genetical evolution of social behaviour, I and II, J. Theoret. Biol., 7, 1—32 (1964).
20. Hamilton W. D., Altruism and related phenomena, mainly in social insects, Ann. Rev. Ecol. Syst, 3, 193—232 (1972).
21. Kettlewell H. B. D. Selection experiments on industrial melanism in the Lepidoptera, Heredity, 10, 14—22 (1956).
22. Kettlewell H. B. D. The Evolution of Melanism, Clarendon Press, Oxford, 1973.
23. Krebs J. R., Davies N. B. An Introduction to Behavioural Ecology, Blackwell Scientific Publications, Oxford, 1981.
24. Lewontin R. C. The units of selection, Ann. Rev. Ecol. Syst., 1, 1—18 (1970).
25. Lewontin R. C. Adaptation, Scientific American, 239, 156—165 (1978).
26. Maynard Smith J. On Evolution, Edinburgh University Press, Edinburgh, 1972.
27. Maynard Smith J. Evolution and the theory of games, American Scientist, 64, 41—45 (1976).
28. Maynard Smith J. The Evolution of Sex, Cambridge University Press, Cambridge, 1978. (Имеется перевод: Мэйнард Смит Дж. Эволюция полового размножения. — М.: Мир, 1981.)
29. Maynard Smith J. Evolution and the Theory of Games, Cambridge University Press, Cambridge, 1982.
30. Metcalf R. A. Sex ratios, parent-offspring conflict, and local competition for mates in the social wasp Polistes metricus, American Naturalist, 116, 642— 654 (1980).
31. Milsum J. H., Roberge F. A. Physiological regulation and control. In: Foundations of Mathematical Biology, 3 (R. Rosen, ed.), Academic Press, London and New York, pp. 1—95, 1973.
32. Muller H. J. The relation of recombination to mutational advance, Mut. Res., 1,2—9 (1954).
33. Parker G. A., Baker R. R., Smith V. G. F. The origin and evolution of gamete dimorphism and the male-female dichotomy, J. Theoret. Biol., 36, 529—553 (1972).
34. Rosen R. Optimality Principles in Biology, Butterworths, London, 1967.
(Имеется перевод: Розен Р. Принципы оптимальности в биологии. — М.:
Мир, 1969.) 35 Sahllns M. The Use and Abuse of Biology: An Anthropological Critique of
Sociobiology, University of Michigan Press, Ann. Arbor, Michigan, 1976.
36. Shorrocks B. The Genesis of Diversity, Hodder and Stoughton, London, 1978.
37. Stearns S. C. Life-history tactics: a review of ideas, Quart. Rev. Biol., 51, 3—47 (1976).
38. Stearns S. C. The evolution of life-history traits: a critique of the theory and a review of the data, Ann. Rev. Ecol.' Syst., 8, 145—171 (1977).
39. Sued J. A., Mayo O. The evolution of dominance. In: Mathematical Topics in Population Genetics (K- Kojima, ed.), Springer Verlag, New York, pp. 289—316, 1970.
40. Townsend C. R., Calow P. (eds.). Physiological Ecology: An evolutionary approach to resource use, Blackwell Scientific Publications, Oxford, 1981.
41. Turner J. R. G. Why does the genome not congeal? Evolution, 21, 645— 656 (1967).
42. Williams G. C. Sex and Evolution, Princeton University Press, Princeton, 1975.
43. Wilson D. S. The Natural Selection of Populations and Communities, Benjamin/Cummings, Menlo Park, California, 1980.
44. Wilson E. 0. Sociobiology: the New Synthesis, Harvard University Press, Harvard, 1975.
45. Wison E. 0., Bossert W. H. A Primer of Population Biology, Sinauer Associates, Stanford, Connecticut, 1971.
Глава 4
1. Alberch P. Ontogenesis and morphological diversification, American Zoologist, 20, 653—667 (1980).
2. Alberch P., Gould S. J., Oster G. F., Wake D. B. Size and shape in ontogeny and phytogeny, Paleobiology, 5, 296—317 (1979).
3. Bard J. B. A unity underlying the different zebra striping patterns, J. Zo-ol. London, 183, 527—539 (1977).
4. Dobzhansky T. Genetics and the Origin of Species, Columbia University Press, New York, 1937.
5. Edmonds H. W., Sawin P. B. Variations of the branches of the aortic arch in rabbits, American Naturalist, 70, 65—66 (1936).
6. Gould S. /. Ontogeny and Phylogeny, Harvard University Press, Cambridge, Mass., 1977.
7. Huxley J. S. Problems of Relative Growth, Methuen, London, 1932.
8. Newell N. D., Phyletic size increase, an important trend illustrated by fossil invertebrates, Evolution, 3, 103—124 (1949).
9. Needham J. On the dissociability of the fundamental processes in ontogenesis, Biol. Revs., 8, 180—223 (1933).
10. Sawin P. В., Edmonds H. W. Morphological studies of the rabbit, VII.
Aortic arch variations in relation to regionally specific growth differences Anat. Rec., 96, 183—200 (1949).
11. Smi/A R. /. Rethinking allometry, J. Theoret. Biol., 87, 97—111 (1980).
12. Waddington С. Н. The Evolution of an Evolutionist, Edinburgh University Press, Edinburgh, 1975,
Глава 5
1. Bus/i G. L. Modes of animal speciation, Ann. Rev. Ecol. Syst., 6, 339—364 (1975).
2. Cronin /. E., Boaz N. Т., Stringer С. В., Rak Y. Tempo and mode in homi-nid evolution, Nature, 292, 113—122 (1981).
3. De Vries H. Species and Varieties. Their Origin by Mutation, The Open Court, Chicago, 1905.
4. Dobzhansky T. Genetics and the Origin of Species, Columbia University Press, New York, 1937.
5. Eldredge N., Cracraft J. Phylogenetic Patterns and the Evolutionary Process, Columbia University Press, New York, 1980.
6. Could S. J., Eldredge N. Punctuated equilibria: the tempo and mode of evolution reconsidered, Paleobiology, 3, 115—151 (1977).
7. Hennig W, Phylogenetic Systematics, 2nd edn., University of Illinois Press, Urbana, 1979.
8. Huxley /. S. Evolution, the Modern Synthesis, Alien and Unwin, London, 1942.
9. Lande R. Natural selection and random genetic drift in phenotypic evolution, 30, 314—334 (1976).
10. Lewin R. Evolution theory under fire, Science, 210, 883—887 (1980).
11. Maynard Smith J. Macroevolution, Nature, 289, 13—14 (1981).
12. Mayr E. Systematics and the Origin of Species, Columbia University Press, New York, 1942. (Имеется перевод: Майр Э. Систематика и происхождение видов. — М.: NG, 1947.)
13. Mayr E. Populations, Species and Evolution, Harvard University Press, Cambridge, Mass.. 1970. (Имеется перевод: Майр Э. Популяции, виды и эволюция. — М.: Мир, 1974.)
14. Mayr E., Provine W. В. The Evolutionary Synthesis, Harvard University Press, Cambridge, Mass., 1980.
15. Simpson G. G. Tempo and Mode in Evolution, Columbia University Press, New York, 1944. (Имеется перевод: Симпсон Дж. Темпы и формы эволюции.—М.: ИЛ, 1948.)
16. Sneath P. H. A., Sokal R. R. Numerical Taxonomy, W. H. Freeman, San Francisco, 1973.
17. Sokal R. R., Sneath P. H. A. Principles of Numerical Taxonomy, W. H. Freeman, San Francisco, 1963.
18. Stanley S. M. At theory of evolution above the species level, Proc Nat Acad. Sci. USA, 72, 646—650 (1975).
19. Stanley S. M. Macroevolution, W. H. Freeman, San Francisco, 1979.
20. Stanley S. M. The New Evolutionary Timetable, Basic Books Inc., New York, 1982.
21. Williams P. G. Palaeontological documentation of speciation in Cenozoic molluscs from Turkana Basin, Nature, 293, 437—443 (1981).